La sensibilidad cutánea comparte los elementos principales de todos los sentidos básicos. Las propiedades del mundo externo, como el color, el sonido o la vibración, son recibidas por terminaciones de células nerviosas especializadas llamadas receptores sensoriales, que convierten los datos externos en impulsos nerviosos. Estas señales luego se transmiten al sistema nervioso central, donde se convierten en la base para interpretar el mundo que nos rodea.
Es útil reconocer tres puntos esenciales sobre estos procesos. Primero, la energía y los cambios en los niveles de energía solo pueden ser percibidos por un órgano sensorial capaz de detectar el tipo específico de energía en cuestión. (Esta es la razón por la cual las microondas, los rayos X y la luz ultravioleta son todos peligrosos; no estamos equipados para detectarlos, por lo que incluso a niveles letales no se perciben). En segundo lugar, nuestras percepciones son necesariamente sombras imperfectas de la realidad, como nuestro centro. sistema nervioso se limita a reconstruir una imagen incompleta a partir de las señales transmitidas por sus receptores sensoriales. En tercer lugar, nuestros sistemas sensoriales nos brindan información más precisa sobre los cambios en nuestro entorno que sobre las condiciones estáticas. Estamos bien equipados con receptores sensoriales sensibles al parpadeo de luces, por ejemplo, oa las mínimas fluctuaciones de temperatura provocadas por una ligera brisa; estamos peor equipados para recibir información sobre una temperatura constante, por ejemplo, o una presión constante sobre la piel.
Tradicionalmente los sentidos de la piel se dividen en dos categorías: cutáneos y profundos. Mientras que la sensibilidad profunda se basa en los receptores ubicados en los músculos, los tendones, las articulaciones y el periostio (membrana que rodea los huesos), la sensibilidad cutánea, que nos ocupa aquí, se ocupa de la información recibida por los receptores en la piel: específicamente, las diversas clases de receptores cutáneos que se encuentran en o cerca de la unión de la dermis y la epidermis.
Todos los nervios sensoriales que unen los receptores cutáneos con el sistema nervioso central tienen aproximadamente la misma estructura. El cuerpo grande de la célula reside en un grupo de otros cuerpos de células nerviosas, llamado ganglio, ubicado cerca de la médula espinal y conectado a ella por una rama estrecha del tronco de la célula, llamada axón. La mayoría de las células nerviosas, o neuronas, que se originan en la médula espinal envían axones a los huesos, músculos, articulaciones o, en el caso de sensibilidad cutánea, a la piel. Al igual que un cable aislado, cada axón está cubierto a lo largo de su curso y en sus extremos con capas protectoras de células conocidas como células de Schwann. Estas células de Schwann producen una sustancia conocida como mielina, que recubre el axón como una vaina. A intervalos a lo largo del camino hay pequeñas rupturas en la mielina, conocidas como nódulos de Ranvier. Finalmente, al final del axón se encuentran los componentes que se especializan en recibir y retransmitir información sobre el ambiente externo: los receptores sensoriales (Mountcastle 1974).
Las diferentes clases de receptores cutáneos, como todos los receptores sensoriales, se definen de dos formas: por sus estructuras anatómicas y por el tipo de señales eléctricas que envían a lo largo de sus fibras nerviosas. Los receptores claramente estructurados suelen llevar el nombre de sus descubridores. Las relativamente pocas clases de receptores sensoriales que se encuentran en la piel se pueden dividir en tres categorías principales: mecanorreceptores, receptores térmicos y nociceptores.
Todos estos receptores pueden transmitir información sobre un estímulo en particular solo después de haberlo codificado primero en un tipo de lenguaje neuronal electroquímico. Estos códigos neuronales utilizan frecuencias y patrones variables de impulsos nerviosos que los científicos apenas han comenzado a descifrar. De hecho, una rama importante de la investigación neurofisiológica se dedica por completo al estudio de los receptores sensoriales y las formas en que traducen los estados de energía del entorno en códigos neuronales. Una vez que se generan los códigos, se transmiten centralmente a lo largo de las fibras aferentes, las células nerviosas que sirven a los receptores al transmitir las señales al sistema nervioso central.
Los mensajes producidos por los receptores se pueden subdividir en función de la respuesta dada a una estimulación continua e invariable: los receptores de adaptación lenta envían impulsos electroquímicos al sistema nervioso central durante la duración de un estímulo constante, mientras que los receptores de adaptación rápida reducen gradualmente sus descargas en la presencia de un estímulo constante hasta que alcanzan un nivel de referencia bajo o cesan por completo, dejando de informar al sistema nervioso central sobre la presencia continua del estímulo.
Las sensaciones claramente diferentes de dolor, calor, frío, presión y vibración son producidas por la actividad en distintas clases de receptores sensoriales y sus fibras nerviosas asociadas. Los términos "aleteo" y "vibración", por ejemplo, se utilizan para distinguir dos sensaciones vibratorias ligeramente diferentes codificadas por dos clases diferentes de receptores sensibles a la vibración (Mountcastle et al. 1967). Las tres categorías importantes de sensación de dolor conocidas como dolor punzante, dolor ardiente y dolor punzante se han asociado cada una con una clase distinta de fibra aferente nociceptiva. Esto no quiere decir, sin embargo, que una sensación específica implique necesariamente sólo una clase de receptor; más de una clase de receptor puede contribuir a una sensación determinada y, de hecho, las sensaciones pueden diferir según la contribución relativa de las diferentes clases de receptor (Sinclair 1981).
El resumen anterior se basa en la hipótesis de especificidad de la función sensorial cutánea, formulada por primera vez por un médico alemán llamado Von Frey en 1906. Aunque durante el siglo pasado se han propuesto al menos otras dos teorías de igual o quizás mayor popularidad, la hipótesis de Von Frey ha ahora ha sido fuertemente respaldada por pruebas fácticas.
Receptores que responden a la presión cutánea constante
En la mano, fibras mielinizadas relativamente grandes (de 5 a 15 mm de diámetro) emergen de una red nerviosa subcutánea llamada plexo nervioso subpapilar y terminan en una ramificación de terminales nerviosas en la unión de la dermis y la epidermis (figura 1). En la piel peluda, estas terminaciones nerviosas culminan en estructuras superficiales visibles conocidas como cúpulas táctiles; en la piel glabra o sin pelo, las terminaciones nerviosas se encuentran en la base de las crestas de la piel (como las que forman las huellas dactilares). Allí, en la cúpula táctil, cada punta de fibra nerviosa, o neurita, está rodeada por una célula epitelial especializada conocida como célula de Merkel (ver figuras 2 y 3).
Figura 1. Una ilustración esquemática de una sección transversal de la piel.
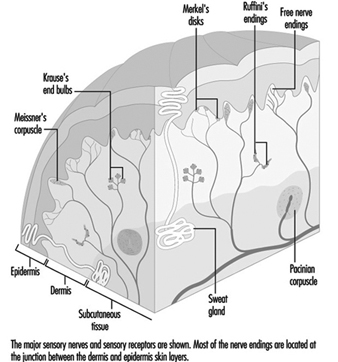
Figura 2. La cúpula táctil en cada región elevada de la piel contiene de 30 a 70 células de Merkel.
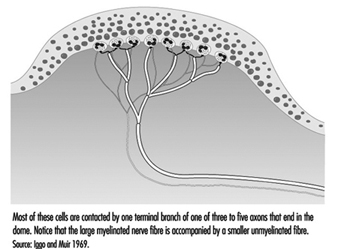
Figura 3. A mayor aumento disponible con el microscopio electrónico, se ve que la célula de Merkel, una célula epitelial especializada, está adherida a la membrana basal que separa la epidermis de la dermis.

El complejo de neuritas de las células de Merkel convierte la energía mecánica en impulsos nerviosos. Si bien se sabe poco sobre el papel de la célula o sobre su mecanismo de transducción, se ha identificado como un receptor de adaptación lenta. Esto significa que la presión sobre una cúpula táctil que contiene células de Merkel hace que los receptores produzcan impulsos nerviosos durante la duración del estímulo. Estos impulsos aumentan de frecuencia en proporción a la intensidad del estímulo, informando así al cerebro de la duración y magnitud de la presión sobre la piel.
Al igual que la célula de Merkel, un segundo receptor de adaptación lenta también sirve a la piel al señalar la magnitud y la duración de las presiones cutáneas constantes. Visible sólo a través de un microscopio, este receptor, conocido como el receptor de Ruffini, consiste en un grupo de neuritas que emergen de una fibra mielinizada y encapsuladas por células de tejido conectivo. Dentro de la estructura de la cápsula hay fibras que aparentemente transmiten distorsiones cutáneas locales a las neuritas, que a su vez producen los mensajes enviados a lo largo de la vía neural al sistema nervioso central. La presión sobre la piel provoca una descarga sostenida de los impulsos nerviosos; al igual que con la célula de Merkel, la frecuencia de los impulsos nerviosos es proporcional a la intensidad del estímulo.
A pesar de sus similitudes, existe una diferencia notable entre las células de Merkel y los receptores de Ruffini. Mientras que la sensación se produce cuando se estimulan los receptores de Ruffini, la estimulación de las cúpulas táctiles que albergan las células de Merkel no produce ninguna sensación consciente; la cúpula táctil es, por lo tanto, un receptor misterioso, ya que se desconoce su papel real en la función neural. Entonces, se cree que los receptores de Ruffini son los únicos receptores capaces de proporcionar las señales neurales necesarias para la experiencia sensorial de presión o contacto constante. Además, se ha demostrado que los receptores de Ruffini que se adaptan lentamente explican la capacidad de los seres humanos para evaluar la presión cutánea en una escala de intensidad.
Receptores que responden a la vibración y al movimiento de la piel
En contraste con los mecanorreceptores de adaptación lenta, los receptores de adaptación rápida permanecen silenciosos durante la indentación sostenida de la piel. Sin embargo, son muy adecuados para señalar la vibración y el movimiento de la piel. Se señalan dos categorías generales: las de piel peluda, que se asocian con pelos individuales; y los que forman terminaciones corpusculares en la piel glabra o sin pelo.
Receptores al servicio de los pelos
Un cabello típico está envuelto por una red de terminales nerviosas que se ramifican de cinco a nueve grandes axones mielinizados (figura 4). En los primates, estas terminales se dividen en tres categorías: terminaciones lanceoladas, terminales fusiformes y terminaciones papilares. Los tres se están adaptando rápidamente, de modo que una desviación constante del cabello provoca impulsos nerviosos solo mientras se produce el movimiento. Por lo tanto, estos receptores son extremadamente sensibles a los estímulos en movimiento o vibratorios, pero brindan poca o ninguna información sobre la presión o el contacto constante.
Figura 4. Los tallos de los cabellos son una plataforma para terminales nerviosas que detectan movimientos.
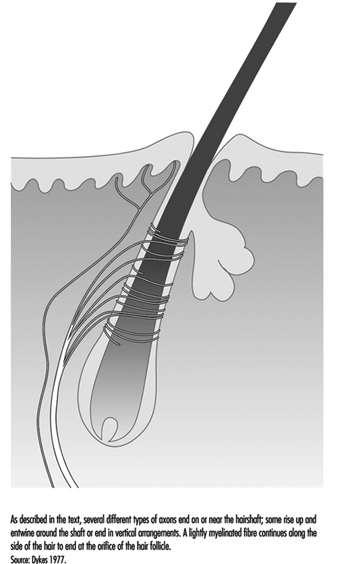
Las terminaciones lanceoladas surgen de una fibra fuertemente mielinizada que forma una red alrededor del cabello. Las neuritas terminales pierden su cobertura habitual de células de Schwann y se abren paso entre las células de la base del cabello.
Los terminales en forma de huso están formados por terminales de axón rodeados por células de Schwann. Los terminales ascienden hasta el tallo del cabello inclinado y terminan en un grupo semicircular justo debajo de una glándula sebácea o productora de aceite. Las terminaciones papilares se diferencian de las terminaciones en forma de huso porque en lugar de terminar en el tallo del cabello, terminan como terminaciones nerviosas libres alrededor del orificio del cabello.
Presumiblemente, existen diferencias funcionales entre los tipos de receptores que se encuentran en los cabellos. Esto se puede deducir en parte de las diferencias estructurales en la forma en que los nervios terminan en el tallo del cabello y en parte de las diferencias en el diámetro de los axones, ya que los axones de diferentes diámetros se conectan a diferentes regiones centrales de transmisión. Aún así, las funciones de los receptores en la piel vellosa siguen siendo un área de estudio.
Receptores en piel glabra
La correlación de la estructura anatómica de un receptor con las señales neurales que genera es más pronunciada en receptores grandes y fácilmente manipulables con terminaciones corpusculares o encapsuladas. Particularmente bien entendidos son los corpúsculos de Pacininan y Meissner, que, como las terminaciones nerviosas en los cabellos discutidos anteriormente, transmiten sensaciones de vibración.
El corpúsculo de Pacini es lo suficientemente grande como para verse a simple vista, lo que facilita vincular el receptor con una respuesta neural específica. Ubicado en la dermis, generalmente alrededor de tendones o articulaciones, es una estructura similar a una cebolla, que mide 0.5 × 1.0 mm. Está servida por una de las fibras aferentes más grandes del cuerpo, que tiene un diámetro de 8 a 13 μm y una velocidad de conducción de 50 a 80 metros por segundo. Su anatomía, bien estudiada tanto por microscopía óptica como electrónica, es bien conocida.
El componente principal del corpúsculo es un núcleo externo formado por material celular que encierra espacios llenos de líquido. El núcleo externo en sí está rodeado por una cápsula que es penetrada por un canal central y una red capilar. A través del canal pasa una sola fibra nerviosa mielínica de 7 a 11 mm de diámetro, que se convierte en una terminación nerviosa larga y no mielinizada que penetra profundamente en el centro del corpúsculo. El axón terminal es elíptico, con procesos ramificados.
El corpúsculo de Pacini es un receptor de rápida adaptación. Cuando se somete a una presión sostenida, produce un impulso solo al principio y al final del estímulo. Responde a vibraciones de alta frecuencia (80 a 400 Hz) y es más sensible a vibraciones alrededor de 250 Hz. A menudo, estos receptores responden a las vibraciones transmitidas a lo largo de los huesos y los tendones y, debido a su extrema sensibilidad, pueden activarse con tan solo una bocanada de aire en la mano (Martin 1985).
Además del corpúsculo de Pacini, hay otro receptor de rápida adaptación en la piel glabra. La mayoría de los investigadores creen que es el corpúsculo de Meissner, ubicado en las papilas dérmicas de la piel. Responde a vibraciones de baja frecuencia de 2 a 40 Hz, este receptor consiste en las ramas terminales de una fibra nerviosa mielinizada de tamaño mediano envuelta en una o varias capas de lo que parecen ser células de Schwann modificadas, llamadas células laminares. Las neuritas y las células laminares del receptor pueden conectarse a una célula basal en la epidermis (figura 5).
Figura 5. El corpúsculo de Meissner es un receptor sensorial débilmente encapsulado en las papilas dérmicas de la piel glabra.
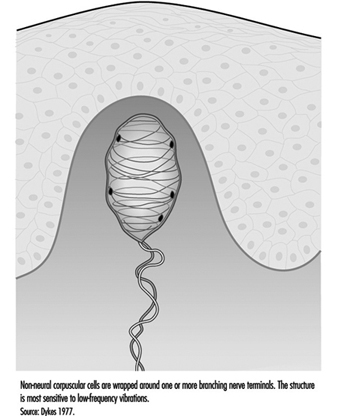
Si el corpúsculo de Meissner se inactiva selectivamente mediante la inyección de un anestésico local a través de la piel, se pierde la sensación de aleteo o vibración de baja frecuencia. Esto sugiere que complementa funcionalmente la capacidad de alta frecuencia de los corpúsculos de Pacini. Juntos, estos dos receptores proporcionan señales neuronales suficientes para explicar la sensibilidad humana a una gama completa de vibraciones (Mountcastle et al. 1967).
Receptores cutáneos asociados con terminaciones nerviosas libres
Muchas fibras mielinizadas y amielínicas aún no identificables se encuentran en la dermis. Un gran número solo está de paso, en su camino hacia la piel, los músculos o el periostio, mientras que otros (tanto mielinizados como no mielinizados) parecen terminar en la dermis. Con unas pocas excepciones, como el corpúsculo de Pacini, la mayoría de las fibras de la dermis parecen terminar en formas mal definidas o simplemente como terminaciones nerviosas libres.
Si bien se necesitan más estudios anatómicos para diferenciar estas terminaciones mal definidas, la investigación fisiológica ha demostrado claramente que estas fibras codifican una variedad de eventos ambientales. Por ejemplo, las terminaciones nerviosas libres que se encuentran en la unión entre la dermis y la epidermis son responsables de codificar los estímulos ambientales que se interpretarán como frío, calor, calor, dolor, picazón y cosquillas. Todavía no se sabe cuál de estas diferentes clases de pequeñas fibras transmite sensaciones particulares.
La aparente similitud anatómica de estas terminaciones nerviosas libres probablemente se deba a las limitaciones de nuestras técnicas de investigación, ya que las diferencias estructurales entre las terminaciones nerviosas libres están saliendo a la luz lentamente. Por ejemplo, en la piel glabra se han distinguido dos modos terminales diferentes de terminaciones nerviosas libres: un patrón grueso y corto y otro largo y delgado. Los estudios de la piel pilosa humana han demostrado terminaciones nerviosas reconocibles histoquímicamente que terminan en la unión dermoepidérmica: las terminaciones penicilada y papilar. Los primeros surgen de fibras amielínicas y forman una red de terminaciones; en cambio, los segundos surgen de fibras mielinizadas y terminan alrededor de los orificios del cabello, como se mencionó anteriormente. Presumiblemente, estas disparidades estructurales corresponden a diferencias funcionales.
Aunque todavía no es posible asignar funciones específicas a entidades estructurales individuales, está claro a partir de experimentos fisiológicos que existen categorías funcionalmente diferentes de terminaciones nerviosas libres. Se ha encontrado que una pequeña fibra mielinizada responde al frío en humanos. Otra fibra amielínica que sirve a las terminaciones nerviosas libres responde al calor. Se desconoce cómo una clase de terminaciones nerviosas libres puede responder selectivamente a una caída de la temperatura, mientras que un aumento de la temperatura de la piel puede provocar que otra clase señale calor. Los estudios muestran que la activación de una fibra pequeña con un extremo libre puede ser responsable de las sensaciones de picazón o cosquilleo, mientras que se cree que hay dos clases de fibras pequeñas específicamente sensibles a los estímulos mecánicos nocivos y químicos o térmicos nocivos, que proporcionan la base neural para los pinchazos. y dolor ardiente (Keele 1964).
La correlación definitiva entre la anatomía y la respuesta fisiológica espera el desarrollo de técnicas más avanzadas. Este es uno de los principales escollos en el manejo de trastornos como la causalgia, la parestesia y la hiperpatía, que siguen siendo un dilema para el médico.
Lesión del nervio periférico
La función neural se puede dividir en dos categorías: sensorial y motora. La lesión de los nervios periféricos, por lo general como resultado del aplastamiento o corte de un nervio, puede afectar la función o ambas, dependiendo de los tipos de fibras en el nervio dañado. Ciertos aspectos de la pérdida motora tienden a malinterpretarse o pasarse por alto, ya que estas señales no van a los músculos sino que afectan el control vascular autónomo, la regulación de la temperatura, la naturaleza y el grosor de la epidermis y el estado de los mecanorreceptores cutáneos. No se discutirá aquí la pérdida de la inervación motora, ni tampoco la pérdida de la inervación que afecta otros sentidos además de los responsables de la sensación cutánea.
La pérdida de la inervación sensorial de la piel crea una vulnerabilidad a más lesiones, ya que deja una superficie anestésica que es incapaz de señalar estímulos potencialmente dañinos. Una vez lesionadas, las superficies de la piel anestesiadas tardan en sanar, quizás en parte debido a la falta de inervación autonómica que normalmente regula factores clave como la regulación de la temperatura y la nutrición celular.
Durante un período de varias semanas, los receptores sensoriales cutáneos desnervados comienzan a atrofiarse, un proceso que es fácil de observar en grandes receptores encapsulados como los corpúsculos de Meissner y Pacini. Si puede ocurrir la regeneración de los axones, puede seguir la recuperación de la función, pero la calidad de la función recuperada dependerá de la naturaleza de la lesión original y de la duración de la denervación (McKinnon y Dellon 1988).
La recuperación después de un aplastamiento de un nervio es más rápida, mucho más completa y más funcional que la recuperación después de que se corta un nervio. Dos factores explican el pronóstico favorable de un aplastamiento de nervios. En primer lugar, es posible que más axones vuelvan a lograr contacto con la piel que después de una transección; segundo, las conexiones son guiadas de vuelta a su sitio original por células de Schwann y revestimientos conocidos como membranas basales, los cuales permanecen intactos en un nervio aplastado, mientras que después de una sección nerviosa, los nervios a menudo viajan a regiones incorrectas de la superficie de la piel siguiendo el trayectorias incorrectas de las células de Schwann. Esta última situación da como resultado que se envíe información espacial distorsionada a la corteza somatosensorial del cerebro. En ambos casos, sin embargo, los axones en regeneración parecen capaces de encontrar el camino de regreso a la misma clase de receptores sensoriales a los que sirvieron anteriormente.
La reinervación de un receptor cutáneo es un proceso gradual. A medida que el axón en crecimiento llega a la superficie de la piel, los campos receptivos son más pequeños de lo normal, mientras que el umbral es más alto. Estos puntos receptivos se expanden con el tiempo y gradualmente se fusionan en campos más grandes. La sensibilidad a los estímulos mecánicos aumenta y, a menudo, se acerca a la sensibilidad de los receptores sensoriales normales de esa clase. Los estudios que utilizan los estímulos del tacto constante, el tacto en movimiento y la vibración han demostrado que las modalidades sensoriales atribuidas a diferentes tipos de receptores regresan a las áreas anestésicas a diferentes velocidades.
Visto bajo un microscopio, la piel glabra denervada se ve más delgada de lo normal, con crestas epidérmicas aplanadas y menos capas de células. Esto confirma que los nervios tienen una influencia trófica o nutricional sobre la piel. Poco después de que vuelve la inervación, las crestas dérmicas se desarrollan mejor, la epidermis se vuelve más gruesa y se pueden encontrar axones que penetran la membrana basal. A medida que el axón regresa al corpúsculo de Meissner, el corpúsculo comienza a aumentar de tamaño y la estructura atrófica previamente aplanada vuelve a su forma original. Si la denervación ha sido de larga duración, se puede formar un nuevo corpúsculo adyacente al esqueleto atrófico original, que permanece denervado (Dellon 1981).
Como puede verse, la comprensión de las consecuencias de la lesión del nervio periférico requiere el conocimiento de la función normal, así como los grados de recuperación funcional. Si bien esta información está disponible para ciertas células nerviosas, otras requieren más investigación, lo que deja una serie de áreas turbias en nuestra comprensión del papel de los nervios cutáneos en la salud y la enfermedad.