皮膚の感受性は、すべての基本的な感覚の主要な要素を共有しています。 色、音、振動などの外界の特性は、感覚受容器と呼ばれる特殊な神経細胞終末によって受信され、外部データを神経インパルスに変換します。 これらの信号は中枢神経系に伝えられ、そこで私たちの周りの世界を解釈するための基礎となります.
これらのプロセスに関する XNUMX つの重要なポイントを認識することは有用です。 まず、エネルギー、およびエネルギー レベルの変化は、問題の特定の種類のエネルギーを検出できる感覚器官によってのみ知覚できます。 (これが、マイクロ波、X 線、紫外線がすべて危険である理由です。私たちはそれらを検出する設備が整っていないため、致死レベルであっても認識されません。) 第二に、私たちの知覚は必然的に現実の不完全な影です。神経系は、その感覚受容体によって伝達される信号から不完全なイメージを再構築することに制限されています。 第三に、私たちの感覚系は、静的な状態よりも環境の変化についてより正確な情報を提供してくれます。 たとえば、ちらつきのある光や、微風によって引き起こされる温度の小さな変動に敏感な感覚受容器が十分に備わっています。 たとえば、一定の温度や皮膚への一定の圧力に関する情報を受け取るには、十分な設備が整っていません。
伝統的に皮膚感覚は、皮膚感覚と深部感覚の XNUMX つのカテゴリーに分けられます。 深い感受性は、筋肉、腱、関節、および骨膜 (骨を囲む膜) にある受容体に依存していますが、ここで関係している皮膚の感受性は、皮膚の受容体によって受信される情報を扱います。具体的には、さまざまなクラスの真皮と表皮の接合部または接合部付近にある皮膚受容体。
皮膚受容体を中枢神経系につなぐすべての感覚神経は、ほぼ同じ構造を持っています。 細胞の大きな体は、神経節と呼ばれる他の神経細胞体のクラスターに存在し、脊髄の近くにあり、軸索と呼ばれる細胞幹の狭い枝によって脊髄に接続されています。 脊髄に由来するほとんどの神経細胞またはニューロンは、軸索を骨、筋肉、関節、または皮膚過敏症の場合は皮膚に送ります。 絶縁されたワイヤーと同じように、各軸索はそのコースに沿って、そしてその終点がシュワン細胞として知られる細胞の保護層で覆われています。 これらのシュワン細胞は、鞘のように軸索を覆うミエリンとして知られる物質を産生します。 ランヴィエ結節として知られるミエリンの小さな断裂が途中で間隔を置いて見られます。 最後に、軸索の末端には、外部環境に関する情報の受信と再送信に特化した構成要素である感覚受容体があります (Mountcastle 1974)。
すべての感覚受容器と同様に、皮膚受容器のさまざまなクラスは、XNUMX つの方法で定義されます。解剖学的構造によるものと、神経線維に沿って送信される電気信号の種類によるものです。 明確な構造の受容体は、通常、発見者にちなんで名付けられます。 皮膚に見られる感覚受容器の比較的少数のクラスは、機械受容器、熱受容器、および侵害受容器の XNUMX つの主要なカテゴリに分類できます。
これらの受容体はすべて、特定の刺激に関する情報を、電気化学的神経言語の一種で最初にエンコードした後にのみ伝達できます。 これらの神経コードは、科学者が解読を始めたばかりの神経インパルスのさまざまな周波数とパターンを使用しています。 実際、神経生理学研究の重要な分野は、感覚受容体と、それらが環境内のエネルギー状態を神経コードに変換する方法の研究に完全に専念しています。 コードが生成されると、信号を中枢神経系に伝達することによって受容体として機能する神経細胞である求心性繊維に沿って中枢に伝達されます。
受容体によって生成されるメッセージは、連続的で不変の刺激に与えられる反応に基づいて細分化できます。ゆっくりと適応する受容体は、一定の刺激の間中枢神経系に電気化学的インパルスを送信しますが、急速に適応する受容体は徐々に放電を減らします。低いベースライン レベルに到達するか、完全に停止するまで安定した刺激が存在し、その後、中枢神経系に刺激の存在が継続していることを通知しなくなります。
痛み、暖かさ、冷たさ、圧力、および振動の明確に異なる感覚は、このように感覚受容器の異なるクラスとそれらに関連する神経線維の活動によって生成されます。 たとえば、「フラッター」と「振動」という用語は、1967 つの異なるクラスの振動感受性受容体によってエンコードされる 1981 つのわずかに異なる振動感覚を区別するために使用されます (Mountcastle et al. XNUMX)。 刺すような痛み、焼けるような痛み、うずくような痛みとして知られる痛覚の XNUMX つの重要なカテゴリは、それぞれ異なるクラスの侵害受容性求心性線維に関連付けられています。 しかし、これは、特定の感覚が必ずしもXNUMXつのクラスの受容体のみを含むと言っているわけではありません。 複数の受容体クラスが特定の感覚に寄与する可能性があり、実際、異なる受容体クラスの相対的な寄与に応じて感覚が異なる場合があります (Sinclair XNUMX)。
前述の要約は、1906 年にフォン フレイという名前のドイツ人医師によって最初に策定された、皮膚感覚機能の特異性仮説に基づいています。過去 XNUMX 世紀の間に、少なくとも XNUMX つの他の同等またはおそらくより人気のある理論が提案されましたが、フォン フレイの仮説は現在、事実に基づく証拠によって強く支持されています。
一定の皮膚圧に反応する受容体
手では、比較的大きな有髄線維 (直径 5 ~ 15 mm) が、乳頭下神経叢と呼ばれる皮下神経網から出て、真皮と表皮の接合部で神経終末のスプレーで終わります (図 1)。 毛むくじゃらの皮膚では、これらの神経終末は、 タッチドーム; 無毛または無毛の皮膚では、神経終末は皮膚隆起の基部 (指紋を形成する隆起など) に見られます。 そこでは、タッチ ドームでは、各神経線維の先端、または神経突起が、 メルケル細胞 (図 2 と 3 を参照)。
図 1. 皮膚の断面の模式図

図 2. 皮膚の隆起した各領域にあるタッチ ドームには、30 ~ 70 のメルケル細胞が含まれています。
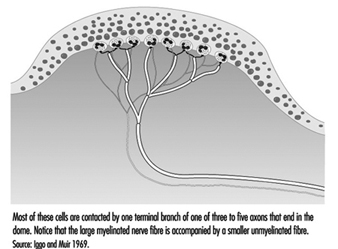
図 3. 電子顕微鏡で利用できる高倍率では、特殊な上皮細胞であるメルケル細胞が、真皮から表皮を分離する基底膜に付着しているのが見られます。
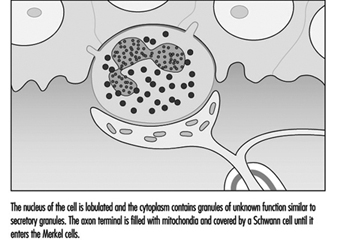
メルケル細胞の神経突起複合体は、機械的エネルギーを神経インパルスに変換します。 細胞の役割や伝達のメカニズムについてはほとんどわかっていませんが、ゆっくりと適応する受容体として同定されています。 これは、メルケル細胞を含むタッチドームへの圧力が、刺激の間、受容体に神経インパルスを生成させることを意味します。 これらのインパルスは、刺激の強さに比例して周波数が上昇し、それによって脳に皮膚への圧力の持続時間と大きさを知らせます。
メルケル細胞と同様に、ゆっくりと適応する XNUMX 番目の受容体も、安定した皮膚圧力の大きさと持続時間を知らせることによって皮膚に働きかけます。 顕微鏡を通してのみ見ることができるこの受容体は、 ルフィニ受容体、有髄繊維から出現し、結合組織細胞によってカプセル化された神経突起のグループで構成されています。 カプセル構造内には、神経突起に局所的な皮膚の歪みを明らかに伝達する繊維があり、神経突起は神経幹線道路に沿って中枢神経系に送信されるメッセージを生成します。 皮膚への圧力は、神経インパルスの持続的な放電を引き起こします。 メルケル細胞と同様に、神経インパルスの頻度は刺激の強さに比例します。
それらの類似性にもかかわらず、メルケル細胞とルフィニ受容体の間には顕著な違いが XNUMX つあります。 ルフィニ受容体が刺激されると感覚が生じるのに対し、メルケル細胞を収容するタッチドームの刺激は意識的な感覚を生じさせません。 したがって、タッチドームは謎の受容体であり、神経機能におけるその実際の役割は不明のままです. したがって、ルフィニ受容体は、圧力や絶え間ない接触の感覚経験に必要な神経信号を提供できる唯一の受容体であると考えられています. さらに、ゆっくりと適応するルフィニ受容体が、強さのスケールで皮膚の圧力を評価する人間の能力を説明することが示されています。
振動と皮膚の動きに反応する受容体
ゆっくりと適応する機械受容体とは対照的に、急速に適応する受容体は持続的な皮膚のインデント中に沈黙を保ちます。 ただし、振動や皮膚の動きの信号には適しています。 XNUMX つの一般的なカテゴリが注目されています。 そして、無毛または無毛の皮膚で粒子末端を形成するもの.
毛髪を処理する受容体
典型的な毛髪は、4 ~ XNUMX 個の大きな有髄軸索から分岐する神経終末のネットワークに包まれています (図 XNUMX)。 霊長類では、これらの終末は、披針形終末、紡錘状終末、乳頭状終末の XNUMX つのカテゴリに分類されます。 XNUMXつすべてが急速に適応しており、動きが発生している間だけ、髪の安定したたわみが神経インパルスを引き起こします. したがって、これらの受容体は、移動または振動刺激に対して非常に敏感ですが、圧力または一定の接触に関する情報はほとんどまたはまったく提供しません.
図 4. 毛幹は、動きを検出する神経終末のプラットフォームです。
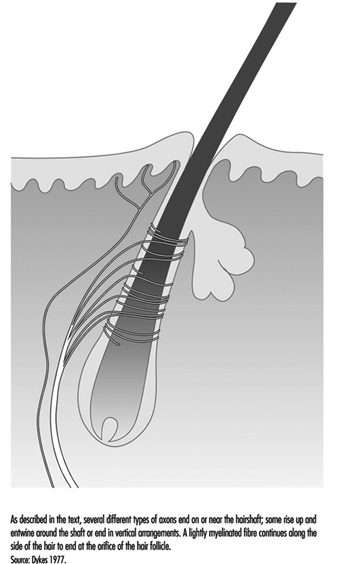
披針形の末端は、髪の周りにネットワークを形成する重度の有髄繊維から生じます。 終末神経突起は、通常のシュワン細胞の被覆を失い、毛の付け根の細胞の間を通り抜けます。
紡錘状終末は、シュワン細胞に囲まれた軸索終末によって形成されます。 端末は、傾斜した毛幹に上昇し、皮脂腺または油産生腺のすぐ下の半円形のクラスターで終わります. 乳頭状終末は紡錘状終末とは異なり、毛幹で終結するのではなく、毛の開口部の周りの自由神経終末として終結します。
おそらく、毛髪に見られる受容体の種類には機能的な違いがあります。 これは、神経が毛幹で終わる方法の構造上の違いと、軸索の直径の違いから部分的に推測できます。これは、異なる直径の軸索が異なる中央中継領域に接続するためです。 それでも、毛むくじゃらの皮膚の受容体の機能は、研究の余地が残されています。
無毛皮膚の受容体
受容体の解剖学的構造と、受容体が生成する神経信号との相関関係は、微粒子またはカプセル化された終末を持つ大きくて操作しやすい受容体で最も顕著です。 特によく理解されているのは、パシニナンとマイスナー小体です。これらは、前述の毛髪の神経終末と同様に、振動の感覚を伝えます。
パチニ小体は肉眼で見えるほど大きいため、受容体と特定の神経反応を簡単に関連付けることができます。 真皮に位置し、通常は腱や関節の周囲にあり、0.5 × 1.0 mm の大きさのタマネギのような構造です。 それは、体の最大の求心性繊維の 8 つで、直径が 13 から 50 μm で、毎秒 80 から XNUMX メートルの速度で伝導しています。 その解剖学は、光学顕微鏡と電子顕微鏡の両方でよく研究されており、よく知られています。
小体の主成分は、液体で満たされた空間を取り囲む細胞物質で形成された外側のコアです。 外核自体は、中心管と毛細血管網が貫通するカプセルに囲まれています。 管を通過するのは、直径 7 ~ 11 mm の XNUMX 本の有髄神経線維であり、小体の中心の奥深くまで突き刺さる長い無髄神経終末になります。 末端軸索は楕円形で、枝のような突起があります。
パチニ小体は急速に適応する受容体です。 したがって、持続的な圧力を受けると、刺激の最初と最後にのみインパルスが生成されます。 高周波振動 (80 ~ 400 Hz) に反応し、250 Hz 付近の振動に最も敏感です。 多くの場合、これらの受容体は骨や腱に沿って伝達される振動に反応し、非常に敏感であるため、手に空気を一吹きするだけで活性化される可能性があります (Martin 1985)。
パチニ小体に加えて、無毛皮膚には急速に適応する受容体がもう 2 つあります。 ほとんどの研究者は、皮膚の真皮乳頭にあるマイスナー小体であると考えています。 40 から 5 Hz の低周波振動に反応するこの受容体は、層状細胞と呼ばれる改変されたシュワン細胞のように見えるものの XNUMX つまたは複数の層に包まれた、中型の有髄神経線維の末端枝で構成されています。 受容体の神経突起と層状細胞は、表皮の基底細胞に接続する可能性があります (図 XNUMX)。
図 5.マイスナー小体は、無毛皮膚の真皮乳頭にゆるくカプセル化された感覚受容体です。
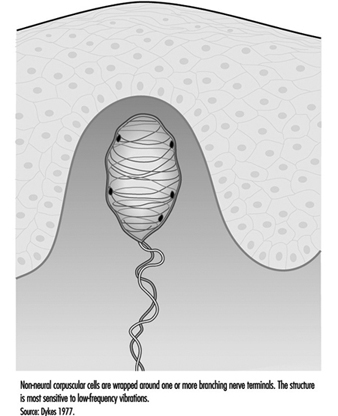
皮膚から局所麻酔薬を注入してマイスナー小体を選択的に不活性化すると、フラッター感や低周波振動が失われます。 これは、パチニ小体の高周波容量を機能的に補完することを示唆しています。 これら 1967 つの受容体は一緒になって、振動の全範囲に対する人間の感受性を説明するのに十分な神経信号を提供します (Mountcastle et al. XNUMX)。
自由神経終末に関連する皮膚受容体
真皮には、まだ識別できない有髄および無髄線維が多く見られます。 多くは、皮膚、筋肉、または骨膜に向かう途中で通過するだけですが、他のもの (有髄および無髄の両方) は真皮で終わるように見えます. パチニ小体などのいくつかの例外を除いて、真皮のほとんどの繊維は、明確に定義されていない方法で終わっているか、単に自由神経終末として終わっているように見えます.
これらの不明確な結末を区別するには、より解剖学的な研究が必要ですが、生理学的研究は、これらの繊維がさまざまな環境イベントをエンコードすることを明確に示しています. たとえば、真皮と表皮の間の接合部に見られる自由神経終末は、寒さ、暖かさ、熱、痛み、かゆみ、およびくすぐりとして解釈される環境刺激をエンコードする役割を果たします。 これらの異なるクラスの小さな繊維のどれが特定の感覚を伝えるかはまだわかっていません.
これらの自由神経終末の明らかな解剖学的類似性は、自由神経終末間の構造上の違いが徐々に明らかになってきているため、おそらく私たちの調査技術の限界によるものです。 たとえば、無毛の皮膚では、自由神経終末の XNUMX つの異なる終末モードが区別されています。太くて短いパターンと長くて細いパターンです。 人間の毛むくじゃらの皮膚の研究は、真皮と表皮の接合部で終結する組織化学的に認識可能な神経終末、すなわちペニシル終末と乳頭終末を示しています。 前者は無髄線維から生じ、終末のネットワークを形成します。 対照的に、後者は有髄繊維から生じ、前述のように毛穴の周りで終わります。 おそらく、これらの構造的不一致は機能的な違いに対応しています。
個々の構造体に特定の機能を割り当てることはまだ不可能ですが、生理学的実験から、自由神経終末には機能的に異なるカテゴリが存在することが明らかです。 小さな有髄繊維の 1964 つが、人間の寒さに反応することがわかっています。 自由な神経終末を提供するもう XNUMX つの無髄線維は、熱に反応します。 自由神経終末の XNUMX つのクラスが温度の低下に選択的に反応し、皮膚温度の上昇が別のクラスを刺激して暖かさを知らせる方法は不明です。 研究によると、自由端を持つ XNUMX つの小さな繊維の活性化がかゆみやくすぐりの感覚の原因である可能性があることが示されていますが、有害な機械的刺激と有害な化学的または熱的刺激に特に敏感な XNUMX つのクラスの小さな繊維があり、刺すための神経基盤を提供すると考えられています。そして焼けるような痛み (Keele XNUMX)。
解剖学と生理学的反応の間の決定的な相関関係は、より高度な技術の開発を待っています。 これは、医師にジレンマを提示し続けている因果痛、知覚異常、および過敏症などの障害の管理における主要な障害の XNUMX つです。
末梢神経損傷
神経機能は、感覚と運動の XNUMX つのカテゴリに分けることができます。 末梢神経損傷は、通常は神経の圧迫または切断に起因し、損傷した神経の繊維の種類に応じて、いずれかまたは両方の機能を損なう可能性があります。 これらの信号は筋肉には伝わらず、むしろ自律血管制御、体温調節、表皮の性質と厚さ、および皮膚の機械受容器の状態に影響を与えるため、運動喪失の特定の側面は誤解されたり見過ごされたりする傾向があります。 運動神経支配の喪失についてはここでは議論しません。また、神経支配の喪失が皮膚感覚以外の感覚に影響を与えることもありません。
皮膚への感覚神経支配の喪失は、潜在的に有害な刺激を伝えることができない麻酔面を残すため、さらなる損傷への脆弱性を生み出します。 いったん損傷すると、麻酔をかけられた皮膚表面は治癒が遅くなります。これはおそらく、体温調節や細胞栄養などの重要な要因を通常は調節する自律神経支配の欠如が原因の XNUMX つです。
数週間にわたって、除神経された皮膚感覚受容体は萎縮し始めます。このプロセスは、パチニ小体やマイスナー小体などのカプセル化された大きな受容体で簡単に観察できます。 軸索の再生が起こる場合、機能の回復が続く可能性がありますが、回復された機能の質は、元の損傷の性質と除神経の期間に依存します (McKinnon and Dellon 1988)。
神経挫傷後の回復は、神経が切断された後の回復よりも迅速で、より完全で機能的です。 神経挫傷の予後が良好であることは、XNUMX つの要因で説明できます。 まず、切断後よりも多くの軸索が再び皮膚との接触を達成する可能性があります。 第二に、接続はシュワン細胞と基底膜として知られる裏打ちによって元の場所に戻されます。これらは両方とも、破壊された神経に無傷のままですが、神経切断後、神経はしばしば皮膚表面の誤った領域に移動します。シュワンセルパスが間違っています。 後者の状況では、歪んだ空間情報が脳の体性感覚皮質に送られます。 しかし、どちらの場合も、再生中の軸索は、以前に機能していたのと同じクラスの感覚受容体に戻る方法を見つけることができるようです。
皮膚受容体の再神経支配は、段階的なプロセスです。 成長中の軸索が皮膚表面に到達すると、受容野は通常より小さくなり、閾値は高くなります。 これらの受容点は時間とともに拡大し、徐々に合体してより大きな領域になります。 機械的刺激に対する感受性はより大きくなり、多くの場合、そのクラスの正常な感覚受容器の感受性に近づきます。 一定の接触、動く接触、および振動の刺激を使用した研究では、さまざまなタイプの受容体に起因する感覚モダリティが、さまざまな速度で麻酔領域に戻ることが示されています。
顕微鏡で見ると、除神経された無毛の皮膚は通常よりも薄く、表皮の隆線が平らになり、細胞の層が少なくなります。 これは、神経が皮膚に栄養的または栄養的な影響を与えることを裏付けています. 神経支配が回復するとすぐに、真皮隆起がより発達し、表皮が厚くなり、基底膜を貫通する軸索が見られます。 軸索がマイスナー小体に戻ると、小体はサイズが大きくなり始め、以前は平らで萎縮した構造が元の形に戻ります。 除神経が長期間続いた場合、除神経されたままの元の萎縮した骨格に隣接して新しい小体が形成される可能性があります (Dellon 1981)。
ご覧のとおり、末梢神経損傷の結果を理解するには、正常な機能と機能回復の程度に関する知識が必要です。 この情報は特定の神経細胞については利用可能ですが、他の細胞についてはさらなる調査が必要であり、健康と病気における皮膚神経の役割を理解するには多くの不明瞭な領域が残されています.