事実上すべての医学は、心筋梗塞、脳卒中、外傷、ショックなどの病気で細胞死を防ぐか、感染症や癌の場合のようにそれを引き起こすことに専念しています. したがって、関与する性質とメカニズムを理解することが不可欠です。 細胞死は、有毒物質や虚血などによって引き起こされる「偶発的」死と、指の形成やオタマジャクシの尾の吸収などの発生過程で発生する「プログラムされた死」に分類されています。
したがって、細胞損傷と細胞死は、生理学と病態生理学の両方で重要です。 生理学的細胞死は、胚形成および胚発生中に非常に重要です。 発生中の細胞死の研究は、特に無脊椎動物の発生の研究を通じて、関連する分子遺伝学に関する重要で新しい情報をもたらしました。 これらの動物では、細胞死を受ける運命にある細胞の正確な位置と重要性が注意深く研究されており、古典的な突然変異誘発技術を使用して、いくつかの関与する遺伝子が現在同定されています. 成人の臓器では、細胞死と細胞増殖のバランスが臓器の大きさを制御します。 皮膚や腸などの一部の臓器では、細胞の絶え間ない代謝回転があります。 例えば皮膚では、細胞は表面に到達するにつれて分化し、架橋エンベロープの形成を伴う角化が進行するにつれて、最終的に最終分化と細胞死を経ます。
有毒化学物質の多くのクラスは、急性の細胞損傷とその後の死を誘発する可能性があります。 これらには、無酸素症と虚血、およびシアン化カリウムなどのそれらの化学的類似体が含まれます。 核酸中のタンパク質に共有結合する求電子剤を形成する化学発癌物質。 酸化剤の化学物質、フリーラジカルの形成と酸化剤の損傷をもたらします。 補体の活性化; およびさまざまなカルシウムイオノフォア。 細胞死は、化学発がんの重要な要素でもあります。 多くの完全な化学発がん物質は、発がん性用量で、急性の壊死と炎症を引き起こし、その後に再生と前腫瘍を引き起こします。
定義
細胞損傷
細胞損傷は、細胞の正常なホメオスタシスを混乱させ、多数のイベントを発生させる有毒化学物質などのイベントまたは刺激として定義されます (図 1)。 説明されている致命的な傷害の主なターゲットは、ATP 合成の阻害、原形質膜の完全性の破壊、または必須の成長因子の離脱です。
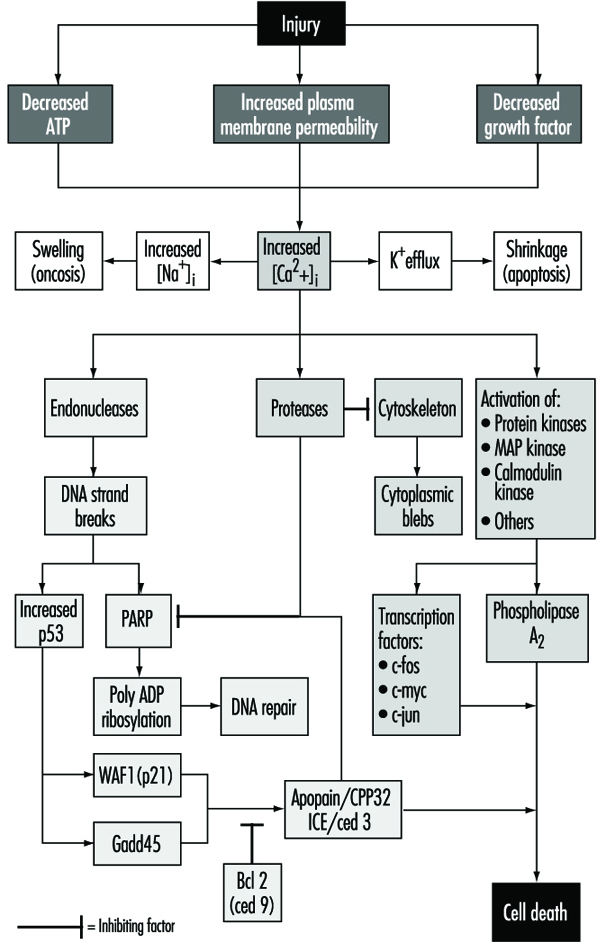
致命的な傷害は、温度、細胞の種類、および刺激に応じて、さまざまな期間の後に細胞の死をもたらします。 またはそれらは致死的または慢性的である可能性があります。つまり、損傷により恒常性状態が変化し、異常ではありますが、細胞死には至りません (Trump and Arstila 1971; Trump and Berezesky 1992; Trump and Berezesky 1995; Trump, Berezesky andオソルニオ・バルガス 1981)。 致命的な損傷の場合、細胞死の前に段階があります
この間、細胞は回復します。 しかし、特定の時点 (「復帰不能点」または細胞死点) の後、損傷の除去は回復には至らず、代わりに細胞は分解と加水分解を受け、最終的に細胞との物理化学的平衡に達します。環境。 これはネクローシスとして知られる段階です。 致死前段階では、細胞と損傷の種類に応じて、いくつかの主要な種類の変化が発生します。 これらは、アポトーシスおよびオンコーシスとして知られています。
アポトーシス
アポトーシスはギリシャ語に由来します アポ、離れていることを意味し、 下垂、落ちることを意味します。 用語 から離れて このタイプの致死前の変化の間に、細胞が収縮し、周辺に顕著な小疱ができるという事実に由来します。 その後、ブレブが剥がれて浮き上がります。 アポトーシスは、さまざまなタイプの毒性損傷に続いて、さまざまな細胞タイプで発生します (Wyllie、Kerr、および Currie 1980)。 これは、リンパ球クローンのターンオーバーの主要なメカニズムであるリンパ球で特に顕著です。 得られた断片は、リンパ節のマクロファージ内に見られる好塩基性体になります。 他の臓器では、アポトーシスは典型的には、隣接する実質細胞またはマクロファージによる断片の食作用によって、死の前後に急速に除去される単一細胞で発生します。 単一細胞で発生するアポトーシスとその後の食作用は、通常、炎症を引き起こしません。 死の前に、アポトーシス細胞は、正常または凝縮したミトコンドリアを持つ非常に高密度のサイトゾルを示します。 小胞体 (ER) は正常であるか、わずかに拡張しています。 核クロマチンは、核膜に沿って核小体の周りに著しく凝集しています。 核の輪郭も不規則で、核の断片化が起こります。 クロマチン凝縮は、多くの場合、ヌクレオソーム間で発生する DNA 断片化に関連しており、電気泳動で特徴的なラダーの外観を示します。
アポトーシスでは、[Ca2+]i Kを刺激する可能性があります+ おそらくATPを必要とする細胞収縮をもたらす流出。 したがって、ATP 合成を完全に阻害する損傷は、アポトーシスを引き起こす可能性が高くなります。 [Caの持続的な増加2+]i プロテアーゼ、エンドヌクレアーゼ、ホスホリパーゼの活性化など、多くの有害な影響があります。 エンドヌクレアーゼの活性化は、一本鎖および二本鎖の DNA 鎖の切断を引き起こし、p53 およびポリ ADP リボシル化、および DNA 修復に不可欠な核タンパク質のレベルの増加を刺激します。 プロテアーゼの活性化は、アクチンおよびブレブ形成につながる関連タンパク質を含む多くの基質を変更します。 もう XNUMX つの重要な基質は、DNA 修復を阻害するポリ (ADP リボース) ポリメラーゼ (PARP) です。 [Caの増加2+]i また、MAPキナーゼ、カルモジュリンキナーゼなどの多くのプロテインキナーゼの活性化にも関連しています。 このようなキナーゼは、c-fos、c-jun、c-myc などの前初期遺伝子の転写を開始する転写因子の活性化、およびホスホリパーゼ A の活性化に関与しています。2 これにより、原形質膜およびミトコンドリアの内膜などの細胞内膜の透過処理が行われます。
腫瘍症
Oncosis、ギリシャ語に由来 ですこのタイプの致死前変化では、損傷の直後に細胞が膨張し始めるため、膨張するという名前が付けられました (Majno and Joris 1995)。 膨潤の理由は、細胞内の水中の陽イオンの増加です。 原因となる主な陽イオンはナトリウムであり、通常は細胞容積を維持するために調節されています。 しかし、ATP が存在しない場合、または原形質膜の Na-ATPase が阻害されている場合、細胞内タンパク質のために体積制御が失われ、水中のナトリウムが増加し続けます。 したがって、腫瘍症の初期のイベントの中で [Na+]i これは細胞の膨張と増加につながります[Ca2+]i 細胞外空間からの流入または細胞内貯蔵からの放出のいずれかによって生じる。 これは、サイトゾルの膨張、小胞体およびゴルジ体の膨張、および細胞表面の周りの水疱の形成をもたらす. ミトコンドリアは最初は凝縮しますが、後にミトコンドリア内膜への損傷により高振幅の膨張を示します。 このタイプの致死前変化では、クロマチンが凝縮し、最終的には分解します。 ただし、アポトーシスの特徴的なはしごパターンは見られません。
壊死
ネクローシスとは、細胞死後、細胞がデブリに変換され、通常は炎症反応によって除去される一連の変化を指します。 腫瘍性壊死とアポトーシス性壊死の XNUMX 種類があります。 浸透圧性壊死は、典型的には、心筋梗塞などの大きなゾーンで、またはHgClの投与後の腎近位尿細管などの化学的毒性の後に臓器の局所的に発生します2. 臓器の広い領域が関与し、壊死細胞が急速に炎症反応を引き起こします。最初は急性で、次に慢性です。 生物が生き残った場合、多くの臓器では、壊死に続いて死んだ細胞が取り除かれ、化学毒性に続いて肝臓や腎臓が再生されます。 対照的に、アポトーシス壊死は通常、単一細胞ベースで発生し、壊死破片はマクロファージまたは隣接する実質細胞の食細胞内で形成されます。 壊死細胞の最も初期の特徴には、原形質膜の連続性の中断と、ミトコンドリアマトリックス内の変性タンパク質を表す凝集密度の出現が含まれます。 最初はミトコンドリアのカルシウム蓄積を妨げない損傷のいくつかの形態では、リン酸カルシウムの沈着物がミトコンドリア内に見られることがあります。 ER、リソソーム、ゴルジ体など、他の膜系も同様に断片化しています。 最終的に、核クロマチンはリソソーム加水分解酵素による攻撃の結果、溶解を受けます。 細胞死に続いて、カテプシン、ヌクレオラーゼ、およびリパーゼを使用して破片を除去する際に、リソソーム加水分解酵素が重要な役割を果たします。これは、これらが最適な酸性 pH を持ち、他の細胞酵素が変性および不活性化されている間、壊死細胞の低 pH を生き残ることができるためです。
メカニズム
初期刺激
致死的な傷害の場合、細胞死につながる傷害をもたらす最も一般的な最初の相互作用は、酸素欠乏症、虚血または呼吸の阻害剤などのエネルギー代謝と、シアン化カリウム、一酸化炭素、ヨード酢酸などの解糖系の干渉です。すぐ。 上述のように、エネルギー代謝を阻害する高用量の化合物は、典型的に腫瘍症を引き起こす。 急性細胞死をもたらす他の一般的なタイプの初期損傷は、原形質膜の機能の改変です (Trump and Arstila 1971; Trump, Berezesky and Osornio-Vargas 1981)。 これは、外傷や補体のC5b-C9複合体の活性化、細胞膜への機械的損傷、またはナトリウム - カリウム(Na+-K+) ウアバインなどのグリコシドをポンプします。 [Ca2+] 勾配を下って細胞に入り、急性致死損傷も引き起こします。 場合によっては、致死前の変化のパターンはアポトーシスです。 他の人では、それは腫瘍症です。
シグナル伝達経路
多くの種類の損傷により、ミトコンドリアの呼吸と酸化的リン酸化が急速に影響を受けます。 一部の細胞では、これは ATP を維持できる嫌気性解糖を刺激しますが、多くの損傷ではこれが阻害されます。 ATP が欠乏すると、多くの重要な恒常性プロセス、特に細胞内イオンの恒常性の制御にエネルギーを与えることができなくなります (Trump and Berezesky 1992; Trump, Berezesky and Osornio-Vargas 1981)。 これにより、[Ca2+]i、および増加した [Na+] および [Cl-] 細胞の膨張を引き起こします。 [Caの増加2+]i 一連のキナーゼを含む、以下で説明する他の多くのシグナル伝達メカニズムの活性化をもたらし、前初期遺伝子転写の増加をもたらす可能性があります。 [Caの増加2+]i また、細胞骨格機能を変更し、部分的に小疱形成を引き起こし、エンドヌクレアーゼ、プロテアーゼ、ホスホリパーゼを活性化します。 これらは、プロテアーゼやリパーゼの活性化による膜損傷、エンドヌクレアーゼの活性化による DNA の直接分解、転写因子として作用する MAP キナーゼやカルモジュリン キナーゼなどのキナーゼの活性化など、上で説明した重要な効果の多くを引き起こすようです。
無脊椎動物の開発に関する広範な研究を通じて C.エレガンス & ショウジョウバエ、ヒトおよび動物の細胞と同様に、一連の死に至る遺伝子が特定されています。 これらの無脊椎動物の遺伝子のいくつかは、哺乳類の対応物を持っていることがわかっています。 たとえば、プログラム細胞死に不可欠な ced-3 遺伝子は、 C. elegans、 プロテアーゼ活性があり、哺乳動物のインターロイキン変換酵素 (ICE) と強い相同性があります。 アポパインまたは prICE と呼ばれる密接に関連する遺伝子が、さらに近い相同性で最近同定されました (Nicholson et al. 1995)。 の ショウジョウバエ、死神遺伝子は、プログラムされた細胞死につながるシグナルに関与しているようです。 他のプロデス遺伝子には、Fas 膜タンパク質と、広く保存されている重要な腫瘍抑制遺伝子 p53 が含まれます。 p53 は、DNA 損傷後にタンパク質レベルで誘導され、リン酸化されると、細胞死シグナル伝達に関与する gadd45 や waf-1 などの他の遺伝子の転写因子として機能します。 c-fos、c-jun、c-myc などの他の前初期遺伝子も、いくつかのシステムに関与しているようです。
同時に、死を促進する遺伝子に対抗するように見える抗死遺伝子があります。 これらのうち最初に特定されたのは、 C.エレガンス、ヒトの bcl-2 と相同です。 これらの遺伝子は、遺伝的毒素または化学的毒素による細胞死を防ぐために、まだ知られていない方法で作用します。 最近のいくつかの証拠は、bcl-2 が抗酸化剤として作用する可能性があることを示しています。 現在、関与する遺伝子の理解を深め、状況に応じてこれらの遺伝子を活性化または阻害する方法を開発するための多くの努力が進行中です.