Praticamente toda a medicina é dedicada a prevenir a morte celular, em doenças como infarto do miocárdio, acidente vascular cerebral, trauma e choque, ou causá-la, como no caso de doenças infecciosas e câncer. É, portanto, essencial entender a natureza e os mecanismos envolvidos. A morte celular tem sido classificada como “acidental”, isto é, causada por agentes tóxicos, isquemia e outros, ou “programada”, como ocorre durante o desenvolvimento embriológico, incluindo formação de dígitos e reabsorção da cauda do girino.
A lesão celular e a morte celular são, portanto, importantes tanto na fisiologia quanto na fisiopatologia. A morte celular fisiológica é extremamente importante durante a embriogênese e o desenvolvimento embrionário. O estudo da morte celular durante o desenvolvimento trouxe importantes e novas informações sobre a genética molecular envolvida, especialmente através do estudo do desenvolvimento em animais invertebrados. Nesses animais, a localização precisa e o significado das células destinadas à morte celular foram cuidadosamente estudados e, com o uso de técnicas clássicas de mutagênese, vários genes envolvidos já foram identificados. Nos órgãos adultos, o equilíbrio entre a morte celular e a proliferação celular controla o tamanho do órgão. Em alguns órgãos, como a pele e o intestino, há uma renovação contínua das células. Na pele, por exemplo, as células se diferenciam à medida que atingem a superfície e, finalmente, sofrem diferenciação terminal e morte celular à medida que a queratinização prossegue com a formação de envelopes reticulados.
Muitas classes de produtos químicos tóxicos são capazes de induzir lesão celular aguda seguida de morte. Estes incluem anóxia e isquemia e seus análogos químicos, como cianeto de potássio; carcinógenos químicos, que formam eletrófilos que se ligam covalentemente a proteínas em ácidos nucléicos; produtos químicos oxidantes, resultando na formação de radicais livres e danos oxidantes; ativação do complemento; e uma variedade de ionóforos de cálcio. A morte celular também é um componente importante da carcinogênese química; muitos carcinógenos químicos completos, em doses carcinogênicas, produzem necrose aguda e inflamação seguida de regeneração e pré-neoplasia.
Definições
lesão celular
A lesão celular é definida como um evento ou estímulo, como um produto químico tóxico, que perturba a homeostase normal da célula, causando assim a ocorrência de vários eventos (figura 1). Os principais alvos de lesão letal ilustrados são a inibição da síntese de ATP, a ruptura da integridade da membrana plasmática ou a retirada de fatores de crescimento essenciais.
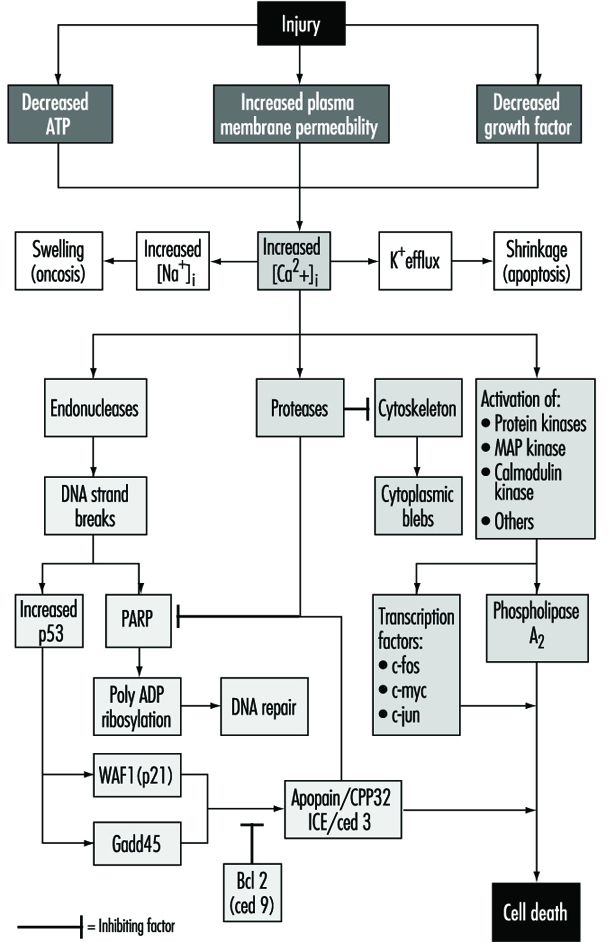
Lesões letais resultam na morte de uma célula após um período de tempo variável, dependendo da temperatura, do tipo de célula e do estímulo; ou podem ser subletais ou crônicos - isto é, a lesão resulta em um estado homeostático alterado que, embora anormal, não resulta em morte celular (Trump e Arstila 1971; Trump e Berezesky 1992; Trump e Berezesky 1995; Trump, Berezesky e Osórnio-Vargas 1981). No caso de uma lesão letal, há uma fase anterior ao momento da morte celular
durante esse tempo, a célula se recuperará; entretanto, após um determinado ponto no tempo (o “ponto sem retorno” ou ponto de morte celular), a remoção da lesão não resulta em recuperação, mas a célula sofre degradação e hidrólise, atingindo finalmente o equilíbrio físico-químico com o meio Ambiente. Esta é a fase conhecida como necrose. Durante a fase pré-letal, vários tipos principais de mudança ocorrem, dependendo da célula e do tipo de lesão. Estes são conhecidos como apoptose e oncose.
Apoptosis
Apoptose é derivado das palavras gregas apo, significando longe de, e ptose, significando cair. O termo caindo longe de deriva do fato de que, durante esse tipo de alteração pré-letal, as células encolhem e sofrem bolhas acentuadas na periferia. As bolhas então se desprendem e flutuam. A apoptose ocorre em uma variedade de tipos de células após vários tipos de lesão tóxica (Wyllie, Kerr e Currie 1980). É especialmente proeminente nos linfócitos, onde é o mecanismo predominante para renovação de clones de linfócitos. Os fragmentos resultantes resultam nos corpos basofílicos vistos dentro dos macrófagos nos gânglios linfáticos. Em outros órgãos, a apoptose ocorre tipicamente em células únicas que são rapidamente eliminadas antes e após a morte por fagocitose dos fragmentos por células parenquimatosas adjacentes ou por macrófagos. A apoptose que ocorre em células únicas com subsequente fagocitose normalmente não resulta em inflamação. Antes da morte, as células apoptóticas apresentam um citosol muito denso com mitocôndrias normais ou condensadas. O retículo endoplasmático (ER) é normal ou apenas ligeiramente dilatado. A cromatina nuclear é marcadamente agrupada ao longo do envelope nuclear e ao redor do nucléolo. O contorno nuclear também é irregular e ocorre fragmentação nuclear. A condensação da cromatina está associada à fragmentação do DNA que, em muitos casos, ocorre entre os nucleossomos, dando uma aparência característica de escada na eletroforese.
Na apoptose, aumentou [Ca2+]i pode estimular K+ efluxo resultando em encolhimento celular, o que provavelmente requer ATP. Lesões que inibem totalmente a síntese de ATP, portanto, têm maior probabilidade de resultar em apoptose. Um aumento sustentado de [Ca2+]i tem uma série de efeitos deletérios, incluindo a ativação de proteases, endonucleases e fosfolipases. A ativação da endonuclease resulta em quebras simples e duplas de DNA que, por sua vez, estimulam níveis aumentados de p53 e na poli-ADP ribosilação, e de proteínas nucleares essenciais no reparo do DNA. A ativação de proteases modifica uma série de substratos, incluindo actina e proteínas relacionadas, levando à formação de bolhas. Outro substrato importante é a poli(ADP-ribose) polimerase (PARP), que inibe o reparo do DNA. Aumentou [Ca2+]i também está associada à ativação de várias proteínas quinases, como MAP quinase, calmodulina quinase e outras. Essas quinases estão envolvidas na ativação de fatores de transcrição que iniciam a transcrição de genes precoces imediatos, por exemplo, c-fos, c-jun e c-myc, e na ativação da fosfolipase A2 que resulta na permeabilização da membrana plasmática e das membranas intracelulares, como a membrana interna da mitocôndria.
oncose
Oncose, derivado da palavra grega É s, inchar, é assim chamado porque neste tipo de alteração pré-letal a célula começa a inchar quase imediatamente após a lesão (Majno e Joris 1995). A razão para o inchaço é um aumento de cátions na água dentro da célula. O principal cátion responsável é o sódio, que normalmente é regulado para manter o volume celular. No entanto, na ausência de ATP ou se a Na-ATPase do plasmalema for inibida, o controle do volume é perdido devido à proteína intracelular e o sódio na água continua a aumentar. Entre os eventos precoces na oncose estão, portanto, o aumento da [Na+]i que leva ao inchaço celular e aumento da [Ca2+]i resultante do influxo do espaço extracelular ou da liberação dos estoques intracelulares. Isso resulta em inchaço do citosol, inchaço do retículo endoplasmático e do aparelho de Golgi e na formação de bolhas aquosas ao redor da superfície celular. As mitocôndrias inicialmente sofrem condensação, mas depois elas também mostram um inchaço de alta amplitude devido a danos na membrana mitocondrial interna. Nesse tipo de alteração pré-letal, a cromatina sofre condensação e, por fim, degradação; no entanto, o padrão de escada característico da apoptose não é observado.
Necrose
Necrose refere-se à série de alterações que ocorrem após a morte celular, quando a célula é convertida em detritos que normalmente são removidos pela resposta inflamatória. Dois tipos podem ser distinguidos: necrose oncótica e necrose apoptótica. A necrose oncótica geralmente ocorre em grandes zonas, por exemplo, em um infarto do miocárdio ou regionalmente em um órgão após toxicidade química, como o túbulo renal proximal após a administração de HgCl2. Amplas zonas de um órgão estão envolvidas e as células necróticas rapidamente incitam uma reação inflamatória, primeiro aguda e depois crônica. No caso de o organismo sobreviver, em muitos órgãos a necrose é seguida pela eliminação das células mortas e regeneração, por exemplo, no fígado ou rim após toxicidade química. Em contraste, a necrose apoptótica ocorre tipicamente em uma única célula e os detritos necróticos são formados dentro dos fagócitos de macrófagos ou células parenquimatosas adjacentes. As primeiras características das células necróticas incluem interrupções na continuidade da membrana plasmática e o aparecimento de densidades floculentas, representando proteínas desnaturadas dentro da matriz mitocondrial. Em algumas formas de lesão que inicialmente não interferem no acúmulo mitocondrial de cálcio, depósitos de fosfato de cálcio podem ser vistos dentro da mitocôndria. Outros sistemas de membrana são fragmentados de forma semelhante, como o RE, os lisossomos e o aparelho de Golgi. Por fim, a cromatina nuclear sofre lise, resultante do ataque das hidrolases lisossômicas. Após a morte celular, as hidrolases lisossômicas desempenham um papel importante na remoção de detritos com catepsinas, nucleolases e lipases, uma vez que estas têm um pH ácido ótimo e podem sobreviver ao baixo pH das células necróticas, enquanto outras enzimas celulares são desnaturadas e inativadas.
Mecanismos
estímulo inicial
No caso de lesões letais, as interações iniciais mais comuns que resultam em lesões que levam à morte celular são a interferência no metabolismo energético, como anoxia, isquemia ou inibidores da respiração, e glicólise, como cianeto de potássio, monóxido de carbono, iodo-acetato e em breve. Como mencionado acima, altas doses de compostos que inibem o metabolismo energético normalmente resultam em oncose. O outro tipo comum de lesão inicial que resulta em morte celular aguda é a modificação da função da membrana plasmática (Trump e Arstila 1971; Trump, Berezesky e Osornio-Vargas 1981). Isso pode ser dano direto e permeabilização, como no caso de trauma ou ativação do complexo C5b-C9 do complemento, dano mecânico à membrana celular ou inibição do sódio-potássio (Na+-K+) bomba com glicosídeos como ouabaína. Ionóforos de cálcio, como ionomicina ou A23187, que transportam rapidamente [Ca2+] descendo o gradiente para dentro da célula, também causam lesões letais agudas. Em alguns casos, o padrão na alteração pré-letal é a apoptose; em outros, é oncose.
Vias de sinalização
Com muitos tipos de lesão, a respiração mitocondrial e a fosforilação oxidativa são rapidamente afetadas. Em algumas células, isso estimula a glicólise anaeróbia, que é capaz de manter o ATP, mas em muitas lesões isso é inibido. A falta de ATP resulta na incapacidade de energizar vários processos homeostáticos importantes, em particular, o controle da homeostase iônica intracelular (Trump e Berezesky 1992; Trump, Berezesky e Osornio-Vargas 1981). Isso resulta em aumentos rápidos de [Ca2+]i, e aumentou [Na+] e [Cl-] resulta em inchaço celular. Aumentos em [Ca2+]i resultam na ativação de vários outros mecanismos de sinalização discutidos abaixo, incluindo uma série de quinases, que podem resultar em aumento imediato da transcrição precoce de genes. Aumentou [Ca2+]i também modifica a função do citoesqueleto, resultando em parte na formação de bolhas e na ativação de endonucleases, proteases e fosfolipases. Estes parecem desencadear muitos dos efeitos importantes discutidos acima, como danos à membrana através da ativação de protease e lipase, degradação direta do DNA pela ativação de endonuclease e ativação de quinases como MAP quinase e calmodulina quinase, que atuam como fatores de transcrição.
Através de um extenso trabalho de desenvolvimento em invertebrados C. elegans e Drosophila, assim como células humanas e animais, uma série de genes pró-morte foram identificados. Descobriu-se que alguns desses genes de invertebrados têm contrapartes de mamíferos. Por exemplo, o gene ced-3, essencial para a morte celular programada em C. elegans, tem atividade de protease e uma forte homologia com a enzima de conversão de interleucina (ICE) de mamíferos. Um gene intimamente relacionado chamado apopaína ou prICE foi recentemente identificado com uma homologia ainda mais estreita (Nicholson et al. 1995). No Drosophila, o gene reaper parece estar envolvido em um sinal que leva à morte celular programada. Outros genes pró-morte incluem a proteína de membrana Fas e o importante gene supressor de tumor, p53, que é amplamente conservado. A p53 é induzida no nível da proteína após o dano ao DNA e quando fosforilada atua como um fator de transcrição para outros genes, como gadd45 e waf-1, que estão envolvidos na sinalização da morte celular. Outros genes precoces imediatos, como c-fos, c-jun e c-myc, também parecem estar envolvidos em alguns sistemas.
Ao mesmo tempo, existem genes anti-morte que parecem neutralizar os genes pró-morte. O primeiro deles a ser identificado foi o ced-9 de C. elegans, que é homólogo ao bcl-2 em humanos. Esses genes agem de uma maneira ainda desconhecida para impedir a morte celular por toxinas genéticas ou químicas. Algumas evidências recentes indicam que o bcl-2 pode atuar como um antioxidante. Atualmente, há muito esforço em andamento para entender os genes envolvidos e desenvolver maneiras de ativar ou inibir esses genes, dependendo da situação.