Сперматогенез и спермиогенез - это клеточные процессы, которые производят зрелые мужские половые клетки. Эти процессы происходят в семенных канальцах семенников половозрелого самца, как показано на рис. 1. Семенные канальцы человека имеют длину от 30 до 70 см и диаметр от 150 до 300 мм (Заневельд, 1978). Сперматогонии (стволовые клетки) расположены вдоль базальной мембраны семенных канальцев и являются основными клетками для производства спермы.
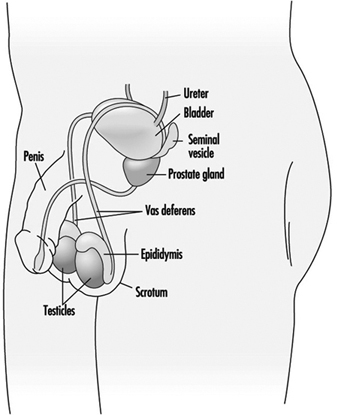
Рисунок 1. Мужская репродуктивная система
Сперматозоиды созревают в результате серии клеточных делений, в которых сперматогонии пролиферируют и становятся первичными сперматоцитами. Покоящиеся первичные сперматоциты мигрируют через плотные соединения, образованные клетками Сертоли, к люминальной стороне этого барьера яичка. К тому времени, когда сперматоциты достигают мембранного барьера в семенниках, синтез ДНК, генетического материала в ядре клетки, практически завершен. Когда первичные сперматоциты действительно попадают в просвет семенных канальцев, они претерпевают особый тип клеточного деления, который происходит только в зародышевых клетках и известен как мейоз. Мейотическое клеточное деление приводит к расщеплению пар хромосом в ядре, так что каждая полученная зародышевая клетка содержит только одну копию каждой цепи хромосомы, а не совпадающую пару.
В ходе мейоза хромосомы изменяют форму, уплотняются и становятся нитевидными. В определенный момент ядерная мембрана, которая их окружает, разрушается, и к хромосомным парам прикрепляются микротрубочковые веретена, вызывая их разделение. На этом завершается первое мейотическое деление и образуются два гаплоидных вторичных сперматоцита. Затем вторичные сперматоциты подвергаются второму мейотическому делению с образованием равного количества сперматид, несущих X- и Y-хромосому.
Морфологическое превращение сперматид в сперматозоиды называется спермиогенезом. Когда спермиогенез завершен, каждая сперматозоид высвобождается клеткой Сертоли в просвет семенных канальцев в результате процесса, называемого спермиацией. Сперматозоиды мигрируют по канальцу к сети яичка и в головку придатка яичка. Сперматозоиды, выходящие из семенных канальцев, незрелые: не могут оплодотворить яйцеклетку и не могут плавать. Сперматозоиды, выбрасываемые в просвет семенных канальцев, находятся во взвешенном состоянии в жидкости, продуцируемой преимущественно клетками Сертоли. Концентрированные сперматозоиды, взвешенные в этой жидкости, непрерывно вытекают из семенных канальцев через небольшие изменения в ионной среде в сети яичка, через эфферентные сосуды и в придаток яичка. Эпидидимис представляет собой единую сильно закрученную трубку (длиной от пяти до шести метров), в которой сперматозоиды проводят от 12 до 21 дня.
В придатке яичка сперматозоиды постепенно приобретают подвижность и способность к оплодотворению. Это может быть связано с изменением характера взвеси жидкости в придатках яичка. То есть по мере созревания клеток придатки яичек поглощают компоненты из жидкости, включая секрецию клеток Сертоли (например, андрогенсвязывающий белок), тем самым увеличивая концентрацию сперматозоидов. Придаток также вносит свой вклад в суспензионную жидкость, включая химические вещества глицерилфосфорилхолин (ГФХ) и карнитин.
Морфология сперматозоидов продолжает трансформироваться в придатках яичка. Цитоплазматическая капля сбрасывается, а ядро спермия еще больше конденсируется. В то время как придатки являются основным резервуаром для хранения спермы до эякуляции, около 30% сперматозоидов в эякуляте хранятся в семявыносящих протоках. Частая эякуляция ускоряет прохождение сперматозоидов через придатки и может увеличить количество незрелых (бесплодных) сперматозоидов в эякуляте (Zaneveld 1978).
эякуляция
Попав в семявыносящие протоки, сперматозоиды транспортируются за счет мышечных сокращений при эякуляции, а не за счет потока жидкости. Во время эякуляции жидкости принудительно выбрасываются из добавочных половых желез, образуя семенную плазму. Эти железы не выделяют свои выделения одновременно. Скорее всего, бульбоуретральная (куперова) железа сначала выделяет прозрачную жидкость, за которой следует секрет предстательной железы, жидкости, концентрированные сперматозоидами, из придатков яичек и ампул семявыносящего протока и, наконец, самая большая фракция, главным образом, из семенных пузырьков. Таким образом, семенная плазма не является однородной жидкостью.
Токсическое действие на сперматогенез и спермиогенез
Токсичные вещества могут нарушить сперматогенез в нескольких точках. Наиболее разрушительными из-за необратимости являются токсиканты, которые убивают или генетически изменяют (без механизмов восстановления) сперматогонии или клетки Сертоли. Исследования на животных были полезны для определения стадии, на которой токсикант воздействует на процесс сперматогенеза. В этих исследованиях используется кратковременное воздействие токсиканта перед отбором проб для определения эффекта. Зная продолжительность каждой стадии сперматогенеза, можно экстраполировать для оценки затронутой стадии.
Биохимический анализ семенной плазмы дает представление о функции добавочных половых желез. Химические вещества, которые секретируются в первую очередь каждой из дополнительных половых желез, обычно выбираются в качестве маркера для каждой соответствующей железы. Например, придатки представлены ГПК, семенные пузырьки — фруктозой, предстательная железа — цинком. Обратите внимание, что этот тип анализа дает только общую информацию о функции желез и мало или совсем не дает информации о других секреторных компонентах. Измерение рН и осмоляльности спермы дает дополнительную общую информацию о природе семенной плазмы.
Семенную плазму можно проанализировать на наличие токсиканта или его метаболита. Тяжелые металлы были обнаружены в семенной плазме с помощью атомно-абсорбционной спектрофотометрии, в то время как галогенированные углеводороды были измерены в семенной жидкости с помощью газовой хроматографии после экстракции или фильтрации с ограничением по белку (Stachel et al., 1989; Zikarge, 1986).
Жизнеспособность и подвижность сперматозоидов в семенной плазме обычно является отражением качества семенной плазмы. Изменения жизнеспособности сперматозоидов, измеренные по эксклюзии окрашивания или гипоосмотическому набуханию, или изменения параметров подвижности сперматозоидов могут свидетельствовать о посттестикулярных токсических эффектах.
Анализы спермы также могут указать, повлиял ли токсикант на выработку сперматозоидов. Количество сперматозоидов и морфология сперматозоидов обеспечивают показатели целостности сперматогенеза и спермиогенеза. Таким образом, количество сперматозоидов в эякуляте прямо коррелирует с количеством половых клеток на грамм яичка (Zukerman et al., 1978), тогда как аномальная морфология, вероятно, является результатом аномального спермиогенеза. Мертвые сперматозоиды или неподвижные сперматозоиды часто отражают последствия посттестикулярных событий. Таким образом, тип или время токсического эффекта может указывать на мишень токсиканта. Например, воздействие 2-метоксиэтанола на самцов крыс привело к снижению фертильности через четыре недели (Chapin et al., 1985). Эти данные, подтвержденные гистологическим исследованием, указывают на то, что мишенью токсичности являются сперматоциты (Chapin et al., 1984). Хотя неэтично преднамеренно подвергать людей воздействию подозреваемых репродуктивных токсикантов, анализы спермы серийных эякулятов мужчин, непреднамеренно подвергшихся воздействию потенциальных токсикантов в течение короткого времени, могут предоставить аналогичную полезную информацию.
Профессиональное воздействие 1,2-дибромхлорпропана (ДБХП) снижало концентрацию сперматозоидов в эякуляте с медианы 79 миллионов клеток/мл у не подвергавшихся воздействию мужчин до 46 миллионов клеток/мл у подвергавшихся воздействию рабочих (Whorton et al., 1979). После удаления рабочих из зоны воздействия у тех, у кого было снижено количество сперматозоидов, произошло частичное выздоровление, в то время как мужчины с азооспермией остались бесплодными. Биопсия яичка показала, что мишенью ДБХП были сперматогонии. Это обосновывает выраженность эффекта, когда мишенью токсикантов являются стволовые клетки. Не было указаний на то, что воздействие ДБХП на мужчин было связано с неблагоприятным исходом беременности (Поташник и Абелиович, 1985). Другим примером токсиканта, воздействующего на сперматогенез/спермогенез, было исследование рабочих, подвергшихся воздействию дибромида этилена (EDB). У них было больше сперматозоидов с заостренными головками и меньше сперматозоидов на эякулят, чем в контрольной группе (Ratcliffe et al., 1987).
Генетическое повреждение трудно обнаружить в сперме человека. Несколько исследований на животных с использованием теста с доминирующей летальностью (Ehling et al., 1978) показывают, что воздействие на отца может привести к неблагоприятному исходу беременности. Эпидемиологические исследования больших групп населения показали повышенную частоту самопроизвольных абортов у женщин, мужья которых работали автомеханиками (McDonald et al., 1989). Такие исследования указывают на необходимость в методах обнаружения генетических повреждений в сперме человека. Такие методы разрабатываются несколькими лабораториями. Эти методы включают зонды ДНК для обнаружения генетических мутаций (Hecht, 1987), кариотипирование хромосом сперматозоидов (Martin, 1983) и оценку стабильности ДНК с помощью проточной цитометрии (Evenson, 1986).
Рисунок 2. Воздействие положительно связано с неблагоприятным влиянием на качество спермы.

На рисунке 2 перечислены воздействия, которые, как известно, влияют на качество спермы, а в таблице 1 представлены сводные результаты эпидемиологических исследований влияния отцов на репродуктивные исходы.
Таблица 1. Эпидемиологические исследования влияния отца на исход беременности
| Справка | Тип воздействия или род занятий | Ассоциация с воздействием1 | эффект |
| Исследования населения на основе записей | |||
| Линдбом и др. 1984 г. | Растворители | – | Самопроизвольный аборт |
| Линдбом и др. 1984 г. | СТО | + | Самопроизвольный аборт |
| Дэниел и Воган, 1988 г. | Органические растворители | – | Самопроизвольный аборт |
| Макдональд и соавт. 1989 | Механика | + | Самопроизвольный аборт |
| Макдональд и соавт. 1989 | Пищевая | + | Дефекты развития |
| Линдбом и др. 1991а | Окись этилена | + | Самопроизвольный аборт |
| Линдбом и др. 1991а | Нефтеперерабатывающий завод | + | Самопроизвольный аборт |
| Линдбом и др. 1991а | Пропитки древесины | + | Самопроизвольный аборт |
| Линдбом и др. 1991а | Резиновые химикаты | + | Самопроизвольный аборт |
| Олсен и др. 1991 г. | Драгоценные металлы | + | Риск детского рака |
| Олсен и др. 1991 г. | Машинисты | + | Риск детского рака |
| Олсен и др. 1991 г. | Кузнецов | + | Риск детского рака |
| Кристенсен и др. 1993 г. | Растворители | + | Преждевременные роды |
| Кристенсен и др. 1993 г. | Свинец и растворители | + | Преждевременные роды |
| Кристенсен и др. 1993 г. | Вести | + | Перинатальная смерть |
| Кристенсен и др. 1993 г. | Вести | + | Заболеваемость детей мужского пола |
| Исследования случай-контроль | |||
| Кучера 1968 г. | Полиграфия | (+) | Заячья губа |
| Кучера 1968 г. | Покраска | (+) | Волчья пасть |
| Олсен 1983 | Покраска | + | Поражение центральной нервной системы |
| Олсен 1983 | Растворители | (+) | Поражение центральной нервной системы |
| Север и др. 1988 г. | Радиация низкого уровня | + | Дефекты нервной трубки |
| Таскинен и др. 1989 г. | Органические растворители | + | Самопроизвольный аборт |
| Таскинен и др. 1989 г. | Ароматические углеводороды | + | Самопроизвольный аборт |
| Таскинен и др. 1989 г. | Пыли | + | Самопроизвольный аборт |
| Гарднер и др. 1990 г. | излучение | + | Детский лейкоз |
| Бонд 1992 | сварка | + | Время до зачатия |
| Уилкинс и Синкс 1990 XNUMX | Сельское хозяйство | (+) | Опухоль головного мозга у ребенка |
| Уилкинс и Синкс 1990 XNUMX | Строительство | (+) | Опухоль головного мозга у ребенка |
| Уилкинс и Синкс 1990 XNUMX | Пищевая/табачная промышленность | (+) | Опухоль головного мозга у ребенка |
| Уилкинс и Синкс 1990 XNUMX | Металл | + | Опухоль головного мозга у ребенка |
| Линдбомн и др. 1991b | Вести | (+) | Самопроизвольный аборт |
| Салмен и др. 1992 г. | Вести | (+) | Врожденные дефекты |
| Веулеманс и др. 1993 г. | Эфир этиленгликоля | + | Аномальная спермограмма |
| Chia et al. 1992 г. | Драгоценные металлы | + | Кадмий в сперме |
1 – нет значимой связи; (+) незначительно значимая ассоциация; + значительная ассоциация.
Источник: адаптировано из Taskinen 1993.
Нейроэндокринная система
Общее функционирование репродуктивной системы контролируется нервной системой и гормонами, вырабатываемыми железами (эндокринная система). Репродуктивная нейроэндокринная ось мужчины включает главным образом центральную нервную систему (ЦНС), переднюю долю гипофиза и яички. Входы от ЦНС и от периферии интегрируются гипоталамусом, который непосредственно регулирует секрецию гонадотропинов передней долей гипофиза. Гонадотропины, в свою очередь, действуют главным образом на клетки Лейдига в интерстиции и клетки Сертоли и зародышевые клетки в семенных канальцах, регулируя сперматогенез и выработку гормонов яичками.
Гипоталамо-гипофизарная ось
Гипоталамус секретирует нейрогормон гонадотропин-рилизинг-гормон (ГнРГ) в портальную сосудистую сеть гипофиза для транспорта в переднюю долю гипофиза. Пульсирующая секреция этого декапептида вызывает сопутствующее высвобождение лютеинизирующего гормона (ЛГ) и, с меньшей синхронностью и в пять раз меньшей силы, высвобождение фолликулостимулирующего гормона (ФСГ) (Bardin, 1986). Существуют существенные доказательства в поддержку наличия отдельного гормона, высвобождающего ФСГ, хотя ни один из них еще не был выделен (Savy-Moore and Schwartz 1980; Culler and Negro-Vilar 1986). Эти гормоны секретируются передней долей гипофиза. ЛГ действует непосредственно на клетки Лейдига, стимулируя синтез и высвобождение тестостерона, тогда как ФСГ стимулирует ароматизацию тестостерона в эстрадиол клетками Сертоли. Гонадотропная стимуляция вызывает высвобождение этих стероидных гормонов в семенные вены.
Секреция гонадотропинов, в свою очередь, контролируется тестостероном и эстрадиолом посредством механизмов отрицательной обратной связи. Тестостерон действует главным образом на гипоталамус, регулируя секрецию ГнРГ и тем самым снижая частоту импульсов, в первую очередь выброса ЛГ. Эстрадиол, с другой стороны, действует на гипофиз, уменьшая величину выброса гонадотропина. Благодаря этим эндокринным петлям обратной связи функция яичек в целом и секреция тестостерона в частности поддерживаются в относительно устойчивом состоянии.
Гипофизарно-тестикулярная ось
ЛГ и ФСГ обычно считаются необходимыми для нормального сперматогенеза. Предположительно эффект ЛГ является вторичным по отношению к индуцированию высокой интратестикулярной концентрации тестостерона. Следовательно, ФСГ из гипофиза и тестостерон из клеток Лейдига воздействуют на клетки Сертоли в эпителии семенных канальцев, инициируя сперматогенез. Продукция сперматозоидов сохраняется, хотя и снижается количественно, после удаления либо ЛГ (и, предположительно, высокой концентрации тестостерона в яичках), либо ФСГ. ФСГ необходим для инициации сперматогенеза в период полового созревания и, в меньшей степени, для возобновления остановленного сперматогенеза (Matsumoto 1989; Sharpe 1989).
Гормональный синергизм, который служит для поддержания сперматогенеза, может повлечь за собой рекрутирование ФСГ дифференцированных сперматогоний для вступления в мейоз, в то время как тестостерон может контролировать конкретные последующие стадии сперматогенеза. ФСГ и тестостерон могут также воздействовать на клетки Сертоли, стимулируя выработку одного или нескольких паракринных факторов, которые могут влиять на количество клеток Лейдига и продукцию тестостерона этими клетками (Sharpe 1989). ФСГ и тестостерон стимулируют синтез белка клетками Сертоли, включая синтез андрогенсвязывающего белка (АСБ), в то время как ФСГ сам по себе стимулирует синтез ароматазы и ингибина. ABP секретируется преимущественно в жидкость семенных канальцев и транспортируется в проксимальную часть головки придатка яичка, возможно, выступая в качестве местного переносчика андрогенов (Bardin 1986). Ароматаза катализирует превращение тестостерона в эстрадиол в клетках Сертоли и других периферических тканях.
Ингибин представляет собой гликопротеин, состоящий из двух несходных субъединиц, связанных дисульфидной связью, а и b. Хотя ингибин предпочтительно ингибирует высвобождение ФСГ, он также может ослаблять высвобождение ЛГ в присутствии стимуляции ГнРГ (Kotsugi et al., 1988). ФСГ и ЛГ стимулируют высвобождение ингибина примерно в равной степени (McLachlan et al., 1988). Интересно, что ингибин секретируется в кровь семенных вен в виде импульсов, синхронных импульсам тестостерона (Winters, 1990). Вероятно, это отражает не прямое действие ЛГ или тестостерона на активность клеток Сертоли, а скорее действие других продуктов клеток Лейдига, секретируемых либо в интерстициальное пространство, либо в кровоток.
Пролактин, который также секретируется передней долей гипофиза, действует синергически с ЛГ и тестостероном, стимулируя мужскую репродуктивную функцию. Пролактин связывается со специфическими рецепторами на клетке Лейдига и увеличивает количество комплекса рецепторов андрогенов в ядрах тканей, чувствительных к андрогенам (Baker et al., 1977). Гиперпролактинемия связана с уменьшением размеров яичек и предстательной железы, объема спермы и концентраций циркулирующего ЛГ и тестостерона (Segal et al., 1979). Гиперпролактинемия также связана с импотенцией, по-видимому, независимо от изменения секреции тестостерона (Thorner et al., 1977).
При измерении метаболитов стероидных гормонов в моче необходимо учитывать вероятность того, что изучаемое воздействие может изменить метаболизм выделяемых метаболитов. Это особенно актуально, поскольку большинство метаболитов образуется в печени, которая является мишенью для многих токсикантов. Свинец, например, уменьшал количество сульфатированных стероидов, выделяемых с мочой (Apostoli et al., 1989). Уровни обоих гонадотропинов в крови повышаются во время сна, когда мужчина достигает половой зрелости, в то время как уровни тестостерона сохраняют этот суточный характер во взрослом возрасте у мужчин (Plant 1988). Таким образом, образцы крови, мочи или слюны следует собирать примерно в одно и то же время суток, чтобы избежать изменений, связанных с суточными секреторными особенностями.
Явные эффекты токсического воздействия на репродуктивную нейроэндокринную систему, скорее всего, проявляются через измененные биологические проявления андрогенов. Проявления, в значительной степени регулируемые андрогенами у взрослого мужчины, которые могут быть обнаружены во время базового медицинского осмотра, включают: (1) задержку азота и развитие мышц; (2) сохранение наружных половых органов и дополнительных половых органов; (3) сохранение увеличенной гортани и утолщенных голосовых связок, обусловливающих мужской голос; (4) рост волос на бороде, в подмышечных впадинах и на лобке, височная рецессия волос и облысение; (5) либидо и сексуальные возможности; (6) органоспецифические белки в тканях (например, в печени, почках, слюнных железах); и (7) агрессивное поведение (Бардин, 1986). Изменения любого из этих признаков могут указывать на нарушение выработки андрогенов.
Примеры токсического воздействия
Свинец — классический пример токсиканта, непосредственно влияющего на нейроэндокринную систему. Концентрации ЛГ в сыворотке были повышены у мужчин, подвергшихся воздействию свинца менее одного года. Этот эффект не прогрессировал у мужчин, подвергавшихся воздействию более пяти лет. Уровень ФСГ в сыворотке не изменился. С другой стороны, уровни АД в сыворотке были повышены, а уровни общего тестостерона снижены у мужчин, подвергшихся воздействию свинца более пяти лет. Уровень свободного тестостерона в сыворотке значительно снизился после воздействия свинца в течение трех-пяти лет (Rodamilans et al., 1988). Напротив, сывороточные концентрации ЛГ, ФСГ, общего тестостерона, пролактина и общего количества нейтральных 17-кетостероидов не изменились у рабочих с более низким уровнем циркулирующего свинца, даже несмотря на то, что частота распределения числа сперматозоидов была изменена (Assennato et al., 1986). .
Воздействие 2-этоксиэтанола на маляров верфи также уменьшало количество сперматозоидов без одновременного изменения концентрации ЛГ, ФСГ или тестостерона в сыворотке (Welch et al., 1988). Таким образом, токсиканты могут независимо влиять на выработку гормонов и показатели спермы.
Рабочие-мужчины, участвующие в производстве нематоцида DBCP, имели повышенные уровни ЛГ и ФСГ в сыворотке крови, а также снижение количества сперматозоидов и снижение фертильности. Эти эффекты, по-видимому, являются следствием действия DBCP на клетки Лейдига с целью изменения продукции или действия андрогенов (Mattison et al., 1990).
Некоторые соединения могут проявлять токсичность в силу структурного сходства с репродуктивными стероидными гормонами. Таким образом, связываясь с соответствующим эндокринным рецептором, токсиканты могут действовать как агонисты или антагонисты, нарушая биологические реакции. Хлордекон (кепон), инсектицид, который связывается с рецепторами эстрогена, снижает количество и подвижность сперматозоидов, останавливает созревание сперматозоидов и снижает либидо. Хотя заманчиво предположить, что эти эффекты являются результатом влияния хлордекона на действие эстрогенов на нейроэндокринном или тестикулярном уровне, в этих исследованиях не было показано, что уровни тестостерона, ЛГ и ФСГ в сыворотке изменились так же, как эффекты терапии эстрадиолом. . ДДТ и его метаболиты также проявляют стероидные свойства и, как можно ожидать, изменяют мужскую репродуктивную функцию, влияя на функции стероидных гормонов. Ксенобиотики, такие как полихлорированные бифенилы, полибромированные бифенилы и хлорорганические пестициды, также могут нарушать репродуктивную функцию мужчин, оказывая агонистическое/антагонистическое действие на эстрогены (Mattison et al., 1990).
Сексуальная функция
Сексуальная функция человека относится к интегрированной деятельности яичек и вторичных половых желез, эндокринной системы контроля и поведенческих и психологических компонентов репродукции (либидо), основанных на центральной нервной системе. Эрекция, эякуляция и оргазм — это три отдельных, независимых, физиологических и психодинамических события, которые обычно происходят одновременно у мужчин.
Имеется мало надежных данных о влиянии профессионального облучения на сексуальную функцию из-за проблем, описанных выше. Было показано, что наркотики влияют на каждую из трех стадий мужской сексуальной функции (Fabro, 1985), что указывает на то, что профессиональные воздействия могут оказывать аналогичные эффекты. Антидепрессанты, антагонисты тестостерона и стимуляторы высвобождения пролактина эффективно снижают либидо у мужчин. Антигипертензивные препараты, действующие на симпатическую нервную систему, у одних мужчин вызывают импотенцию, а у других — приапизм. Феноксибензамин, антагонист адренорецепторов, использовался в клинических условиях для блокирования выброса семени, но не оргазма (Shilon, Paz and Homonnai, 1984). Антихолинергические антидепрессанты позволяют семяизвержение, блокируя выброс семенной жидкости и оргазм, что приводит к просачиванию семенной плазмы из уретры, а не выбросу.
Рекреационные наркотики также влияют на сексуальную функцию (Fabro, 1985). Этанол может уменьшить импотенцию, одновременно повышая либидо. Кокаин, героин и высокие дозы каннабиноидов снижают либидо. Опиаты также задерживают или ухудшают эякуляцию.
Огромный и разнообразный спектр фармацевтических препаратов, которые, как было доказано, влияют на мужскую репродуктивную систему, подтверждает идею о том, что химические вещества, обнаруженные на рабочем месте, также могут быть репродуктивными токсичными веществами. Для оценки этой важной области репродуктивной токсикологии необходимы надежные и практичные методы исследования для условий полевых исследований.