Рисунок 1. Женская репродуктивная система.
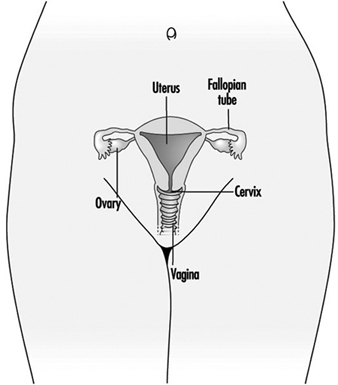
Женская репродуктивная система контролируется компонентами центральной нервной системы, включая гипоталамус и гипофиз. Она состоит из яичников, фаллопиевых труб, матки и влагалища (рис. 1). Яичники, женские гонады, являются источником ооцитов, а также синтезируют и секретируют эстрогены и прогестагены, основные женские половые гормоны. Фаллопиевы трубы транспортируют ооциты и сперму из матки. Матка представляет собой грушевидный мышечный орган, верхняя часть которого сообщается через фаллопиевы трубы с брюшной полостью, а нижняя — через узкий канал шейки матки соприкасается с влагалищем, выходящим наружу. В таблице 1 приведены соединения, клинические проявления, места и механизмы действия потенциальных репродуктивных токсикантов.
Таблица 1. Потенциальные токсиканты женской репродуктивной системы
| Соединение | Клиническое проявление | Сайт | Механизм/мишень |
| Химическая реактивность | |||
| алкилирующий агенты |
Измененные менструации Аменорея атрофия яичников Снижение фертильности Преждевременная менопауза |
яичник матка |
Цитотоксичность гранулезных клеток Цитотоксичность ооцитов Цитотоксичность клеток эндометрия |
| Вести | Аномальные менструации атрофия яичников Снижение фертильности |
Гипоталамус Гипофиз яичник |
Снижение ФСГ Снижение прогестерона |
| ртутный | Аномальные менструации | Гипоталамус яичник |
Изменение продукции и секреции гонадотропинов Фолликулярная токсичность Пролиферация гранулезных клеток |
| Кадмий | Фолликулярная атрезия Стойкий диэструс |
яичник Гипофиз Гипоталамус |
Сосудистая токсичность Цитотоксичность гранулезных клеток цитотоксичность |
| Структурное сходство | |||
| Азатиоприн | Уменьшенное количество фолликулов | яичник оогенеза |
Аналог пурина Нарушение синтеза ДНК/РНК |
| Хлордекон | Нарушение фертильности | Гипоталамус | Агонист эстрогена |
| ДДТ | Измененные менструации | Гипофиз | нарушение ФСГ, ЛГ |
| 2,4-D | бесплодность | ||
| Lindane | Аменорея | ||
| Токсафен | Гиперменорея | ||
| ПХД, ПБД | Аномальные менструации | нарушение ФСГ, ЛГ | |
Источник: Из Plowchalk, Meadows and Mattison 1992. Предполагается, что эти соединения являются репродуктивными токсикантами прямого действия, основываясь главным образом на тестах токсичности на экспериментальных животных.
Гипоталамус и гипофиз
Гипоталамус расположен в промежуточном мозге, который находится на вершине ствола мозга и окружен полушариями головного мозга. Гипоталамус является основным посредником между нервной и эндокринной системами, двумя главными системами управления тела. Гипоталамус регулирует работу гипофиза и выработку гормонов.
Механизмы, с помощью которых химическое вещество может нарушить репродуктивную функцию гипоталамуса, обычно включают любое событие, которое может изменить пульсирующее высвобождение гонадотропин-высвобождающего гормона (ГнРГ). Это может включать изменение либо частоты, либо амплитуды импульсов ГнРГ. Процессы, подверженные химическому повреждению, включают процессы, участвующие в синтезе и секреции GnRH, в частности, транскрипцию или трансляцию, упаковку или аксональный транспорт и секреторные механизмы. Эти процессы представляют собой места, где химически активные соединения прямого действия могут мешать гипоталамическому синтезу или высвобождению GnRH. Изменение частоты или амплитуды импульсов ГнРГ может быть результатом нарушений в стимулирующих или ингибирующих путях, которые регулируют высвобождение ГнРГ. Исследования регуляции генератора импульсов ГнРГ показали, что катехоламины, дофамин, серотонин, γ-аминомасляная кислота и эндорфины обладают определенным потенциалом для изменения высвобождения ГнРГ. Следовательно, ксенобиотики, являющиеся агонистами или антагонистами этих соединений, могут модифицировать высвобождение ГнРГ, тем самым нарушая связь с гипофизом.
Пролактин, фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ) представляют собой три белковых гормона, секретируемых передней долей гипофиза, которые необходимы для репродукции. Они играют критическую роль в поддержании овариального цикла, контролируя рекрутирование и созревание фолликулов, стероидогенез, завершение созревания яйцеклеток, овуляцию и лютеинизацию.
Точный, точно настроенный контроль репродуктивной системы осуществляется передней долей гипофиза в ответ на положительные и отрицательные сигналы обратной связи от половых желез. Надлежащий выброс ФСГ и ЛГ во время овариального цикла контролирует нормальное развитие фолликулов, а отсутствие этих гормонов сопровождается аменореей и атрофией гонад. Гонадотропины играют критическую роль в инициировании изменений в морфологии фолликулов яичников и в их стероидном микроокружении посредством стимуляции продукции стероидов и индукции популяций рецепторов. Своевременное и адекватное высвобождение этих гонадотропинов также важно для овуляторных событий и функциональной лютеиновой фазы. Поскольку гонадотропины необходимы для функции яичников, измененный синтез, накопление или секреция могут серьезно нарушить репродуктивную способность. Вмешательство в экспрессию генов — будь то транскрипция или трансляция, посттрансляционные события или упаковка или секреторные механизмы — может изменить уровень гонадотропинов, достигающих половых желез. Химические вещества, которые действуют за счет структурного сходства или изменения эндокринного гомеостаза, могут вызывать эффекты, вмешиваясь в нормальные механизмы обратной связи. Агонисты и антагонисты стероидных рецепторов могут инициировать неадекватное высвобождение гонадотропинов из гипофиза, тем самым индуцируя ферменты, метаболизирующие стероиды, снижая период полувыведения стероидов и, следовательно, уровень циркулирующих стероидов, достигающих гипофиза.
яичник
Яичник у приматов отвечает за контроль размножения посредством своих основных продуктов, ооцитов, стероидных и белковых гормонов. Фолликулогенез, в котором задействованы как интраовариальные, так и экстраовариальные регуляторные механизмы, представляет собой процесс, посредством которого вырабатываются ооциты и гормоны. Сам яичник состоит из трех функциональных субъединиц: фолликула, ооцита и желтого тела. Во время нормального менструального цикла эти компоненты, под влиянием ФСГ и ЛГ, функционируют согласованно, создавая жизнеспособную яйцеклетку для оплодотворения и подходящую среду для имплантации и последующей беременности.
В преовуляторный период менструального цикла рекрутирование и развитие фолликулов происходит под влиянием ФСГ и ЛГ. Последний стимулирует выработку андрогенов тека-клетками, тогда как первый стимулирует ароматизацию андрогенов в эстрогены гранулезными клетками и выработку ингибина, белкового гормона. Ингибин действует на переднюю долю гипофиза, уменьшая высвобождение ФСГ. Это предотвращает избыточную стимуляцию развития фолликулов и позволяет продолжить развитие доминантного фолликула — фолликула, которому суждено овулировать. Выработка эстрогена увеличивается, стимулируя как всплеск ЛГ (что приводит к овуляции), так и клеточные и секреторные изменения во влагалище, шейке матки, матке и яйцеводе, которые повышают жизнеспособность и транспорт сперматозоидов.
В постовуляторную фазу тека- и гранулезные клетки, остающиеся в фолликулярной полости овулировавшей яйцеклетки, образуют желтое тело и секретируют прогестерон. Этот гормон стимулирует матку, чтобы обеспечить надлежащую среду для имплантации эмбриона, если происходит оплодотворение. В отличие от мужской гонады, женские гонады имеют конечное число зародышевых клеток при рождении и поэтому обладают уникальной чувствительностью к репродуктивным токсикантам. Такое воздействие на самку может привести к снижению плодовитости, увеличению невынашивания беременности, ранней менопаузе или бесплодию.
Являясь основной репродуктивной единицей яичника, фолликул поддерживает деликатную гормональную среду, необходимую для поддержки роста и созревания ооцита. Как отмечалось ранее, этот сложный процесс известен как фолликулогенез и включает как интраовариальную, так и экстраовариальную регуляцию. Многочисленные морфологические и биохимические изменения происходят по мере того, как примордиальный фолликул прогрессирует до преовуляторного фолликула (который содержит развивающийся ооцит), и каждая стадия роста фолликула демонстрирует уникальные модели чувствительности к гонадотропинам, продукции стероидов и путей обратной связи. Эти характеристики позволяют предположить, что ряд сайтов доступен для взаимодействия ксенобиотиков. Кроме того, в яичниках имеются разные популяции фолликулов, что еще больше усложняет ситуацию, допуская дифференциальную токсичность фолликулов. Это создает ситуацию, в которой модели бесплодия, вызванные химическим агентом, будут зависеть от типа пораженного фолликула. Например, токсичность для примордиальных фолликулов не вызовет немедленных признаков бесплодия, но в конечном итоге сократит репродуктивную продолжительность жизни. С другой стороны, токсичность для антральных или преовуляторных фолликулов может привести к немедленной потере репродуктивной функции. Фолликулярный комплекс состоит из трех основных компонентов: гранулезных клеток, тека-клеток и ооцита. Каждый из этих компонентов обладает характеристиками, которые могут сделать его уникально восприимчивым к химическому повреждению.
Несколько исследователей изучили методологию скрининга ксенобиотиков на токсичность клеток гранулезы путем измерения влияния на выработку прогестерона клетками гранулезы в культуре. Подавление эстрадиолом выработки прогестерона клетками гранулезы использовалось для проверки чувствительности клеток гранулезы. Пестицид p,p'-ДДТ и его изомер o,p'-ДДТ подавляют выработку прогестерона, по-видимому, с эффективностью, равной эстрадиолу. Напротив, пестициды малатион, аратион и дильдрин, а также фунгицид гексахлорбензол не действуют. Необходим дальнейший подробный анализ ответов изолированных клеток гранулезы на ксенобиотики, чтобы определить полезность этой системы анализа. Привлекательность таких изолированных систем заключается в экономичности и простоте использования; однако важно помнить, что клетки гранулезы представляют собой только один компонент репродуктивной системы.
Текальные клетки являются предшественниками стероидов, синтезируемых клетками гранулезы. Текальные клетки, как полагают, рекрутируются из клеток стромы яичников во время образования и роста фолликулов. Рекрутинг может включать пролиферацию стромальных клеток, а также миграцию в области вокруг фолликула. Ксенобиотики, которые нарушают пролиферацию, миграцию и коммуникацию клеток, будут влиять на функцию текальных клеток. Ксенобиотики, которые изменяют продукцию андрогенов в тканях, также могут нарушать функцию фолликулов. Например, андрогены, метаболизируемые клетками гранулезы в эстрогены, поступают из тека-клеток. Ожидается, что изменения в продукции андрогенов текальными клетками, будь то увеличение или уменьшение, окажут значительное влияние на функцию фолликула. Например, считается, что избыточная продукция андрогенов клетками теки приводит к атрезии фолликулов. Кроме того, нарушение продукции андрогенов тека-клетками может привести к снижению продукции поэстрогенов гранулезными клетками. Любое обстоятельство явно повлияет на репродуктивную функцию. В настоящее время мало что известно об уязвимости текальных клеток к ксенобиотикам.
Хотя имеется достаточно информации, определяющей уязвимость клеток яичников к ксенобиотикам, имеются данные, ясно демонстрирующие, что ооциты могут быть повреждены или разрушены такими агентами. Алкилирующие агенты разрушают ооциты человека и экспериментальных животных. Свинец оказывает токсическое действие на яичники. Ртуть и кадмий также вызывают повреждение яичников, которое может быть опосредовано через токсичность ооцитов.
От оплодотворения до имплантации
Гаметогенез, высвобождение и объединение мужских и женских половых клеток — все это предварительные события, ведущие к зиготе. Сперматозоиды, депонированные во влагалище, должны попасть в шейку матки и пройти через матку в маточную трубу, чтобы встретиться с яйцеклеткой. проникновение сперматозоидов в яйцеклетку и слияние соответствующих ДНК составляют процесс оплодотворения. После оплодотворения начинается деление клеток, которое продолжается в течение следующих трех или четырех дней, образуя твердую массу клеток, называемую морулой. Клетки морулы продолжают делиться, и к тому времени, когда развивающийся эмбрион достигает матки, он представляет собой полый шар, называемый бластоцистой.
После оплодотворения развивающийся эмбрион мигрирует по фаллопиевой трубе в матку. Бластоциста попадает в матку и имплантируется в эндометрий примерно через семь дней после овуляции. В это время эндометрий находится в постовуляторной фазе. Имплантация позволяет бластоцисте поглощать питательные вещества или токсиканты из желез и кровеносных сосудов эндометрия.