人們早就認識到,每個人對環境化學品的反應是不同的。 最近分子生物學和遺傳學的爆炸式增長使人們對這種變異性的分子基礎有了更清晰的認識。 個體對化學品反應的主要決定因素包括十幾個酶超家族之間的重要差異,統稱為 異生素- (身體異物)或 藥物代謝 酵素。 雖然這些酶的作用傳統上被認為是解毒,但這些相同的酶也會將許多惰性化合物轉化為劇毒中間體。 最近,已經確定了編碼這些酶的基因的許多細微和明顯的差異,這些差異已被證明會導致酶活性的顯著變化。 現在很清楚,每個個體都擁有獨特的異生素代謝酶活性; 這種多樣性可能被認為是“代謝指紋”。 正是這些許多不同的酶超家族之間複雜的相互作用最終不僅決定了化學品在任何給定個體中的命運和潛在毒性,而且還決定了對暴露的評估。 在這篇文章中,我們選擇使用細胞色素 P450 酶超家族來說明在理解個體對化學品的反應方面取得的顯著進步。 旨在識別這些酶中特定基因改變的相對簡單的基於 DNA 的測試的開發,現在可以更準確地預測個體對化學暴露的反應。 我們希望結果將是預防性毒理學。 換句話說,每個人都可以了解他或她特別敏感的那些化學物質,從而避免以前無法預測的毒性或癌症。
儘管人們普遍不理解,但人類每天都會接觸到無數種不同的化學物質。 這些化學物質中有許多是劇毒的,它們來自各種各樣的環境和飲食來源。 這種暴露與人類健康之間的關係一直並將繼續成為全球生物醫學研究工作的主要焦點。
這種化學轟擊有哪些例子? 超過 400 種來自紅酒的化學物質已被分離和表徵。 據估計,一根點燃的香煙至少會產生 1,000 種化學物質。 化妝品和香皂中含有無數化學物質。 接觸化學品的另一個主要來源是農業:僅在美國,農田每年就會收到超過 75,000 種農藥、除草劑和肥料形式的化學品; 在被植物和食草動物以及附近水道中的魚類吸收後,人類(處於食物鏈的末端)攝入這些化學物質。 攝入體內高濃度化學物質的其他兩個來源包括 (a) 長期服用藥物和 (b) 在整個職業生涯中接觸工作場所的有害物質。
現在已經確定,化學品接觸可能對人類健康的許多方面產生不利影響,導致慢性疾病和許多癌症的發展。 在過去十年左右的時間裡,許多這些關係的分子基礎已經開始被闡明。 此外,人們已經認識到,人類對接觸化學品有害影響的敏感性存在顯著差異。
目前預測人類對化學暴露反應的努力結合了兩種基本方法(圖 1):通過生物標記物(生物標誌物)監測人類暴露的程度,以及預測個體對給定暴露水平的可能反應。 儘管這兩種方法都非常重要,但應該強調的是,這兩種方法截然不同。 本文將重點介紹 遺傳因素 潛在的個體對任何特定化學品暴露的易感性。 這個研究領域被廣泛稱為 生態遺傳學, 或者 藥物遺傳學 (參見 Kalow 1962 和 1992)。 許多最近在確定個體對化學毒性的易感性方面的進展源於對人類和其他哺乳動物對化學物質解毒的過程以及所涉及的酶系統的顯著複雜性的更深入了解。
圖 1. 暴露評估、種族差異、年齡、飲食、營養和遺傳易感性評估之間的相互關係——所有這些都在個體中毒和癌症風險中發揮作用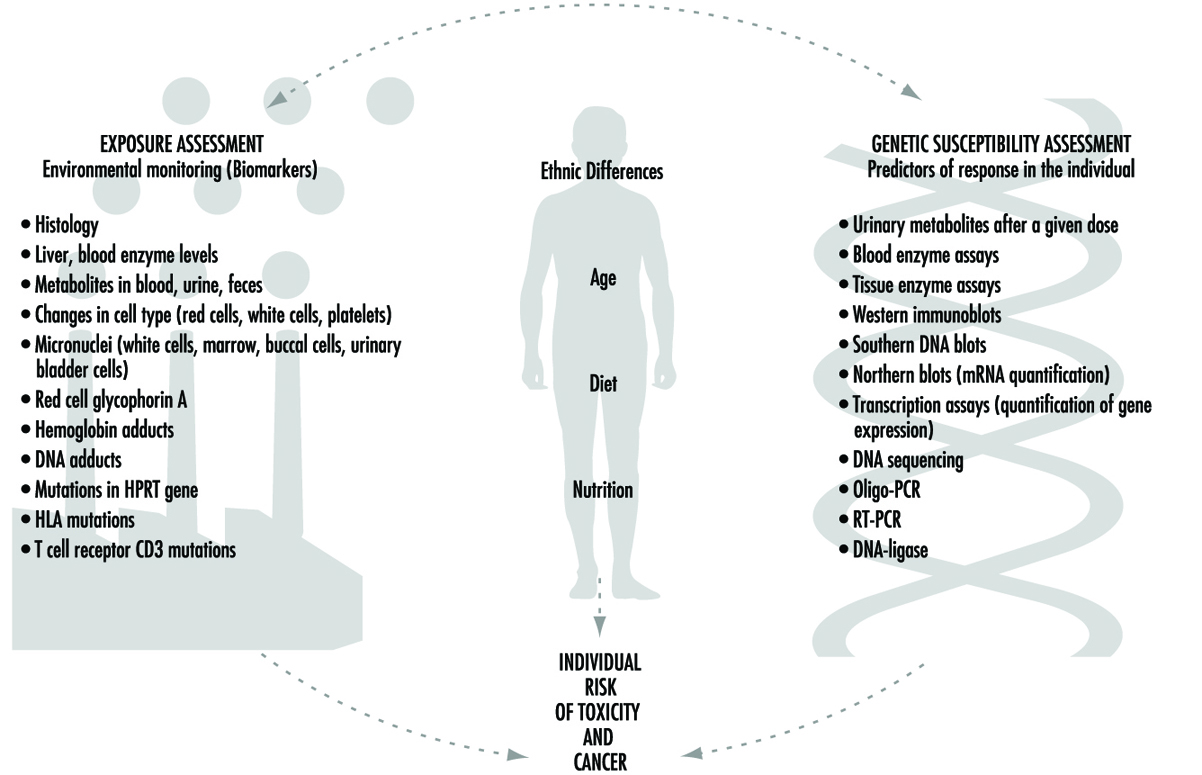
我們將首先描述人類毒性反應的可變性。 然後,我們將介紹一些由於外來化學物質的代謝差異而導致響應變化的酶。 接下來,將詳細介紹細胞色素 P450 超家族的歷史和命名法。 將簡要描述五種人類 P450 多態性以及幾種非 P450 多態性; 這些是造成人類毒性反應差異的原因。 然後我們將討論一個例子來強調這一點,即個體的遺傳差異會影響暴露評估,這是由環境監測確定的。 最後,我們將討論這些異生素代謝酶在關鍵生命功能中的作用。
人群中毒性反應的變化
毒理學家和藥理學家通常談論 50% 人口的平均致死劑量 (LD50), 50% 人群的平均最大耐受劑量 (MTD50),以及特定藥物對 50% 人口的平均有效劑量 (ED50). 然而,這些劑量如何影響我們每個人的個人基礎? 換句話說,一個高度敏感的人可能比人群中抵抗力最強的人受影響 500 倍或受影響的可能性高 500 倍; 對於這些人,LD50 (和MTD50 和艾德50) 值沒有什麼意義。 低密度脂蛋白50, 最大傳輸距離50 和艾德50 只有在提及整個人口時,價值觀才有意義。
圖2 說明了任何給定人群中個體對毒性反應的假設劑量反應關係。 這個通用圖表可能表示支氣管癌與吸煙數量的關係,氯痤瘡與工作場所二噁英水平的關係,哮喘與空氣中臭氧或醛濃度的關係,曬傷與紫外線的關係,凝血時間縮短作為阿司匹林攝入量的函數,或胃腸道不適對服用次數的反應 辣椒 辣椒消耗。 通常,在這些情況中的每一種情況下,接觸越多,毒性反應就越大。 大多數人群將表現出作為劑量函數的毒性反應的平均值和標準偏差。 “耐藥異常值”(圖 2 右下角)是指在較高劑量或暴露條件下反應較少的個體。 “敏感異常值”(左上角)是對相對較小的劑量或暴露有誇大反應的個體。 這些異常值與人群中的大多數個體相比反應存在極大差異,可能代表重要的遺傳變異,可以幫助科學家試圖了解毒性反應的潛在分子機制。
圖 2. 任何毒性反應與任何環境、化學或物理因素劑量之間的一般關係
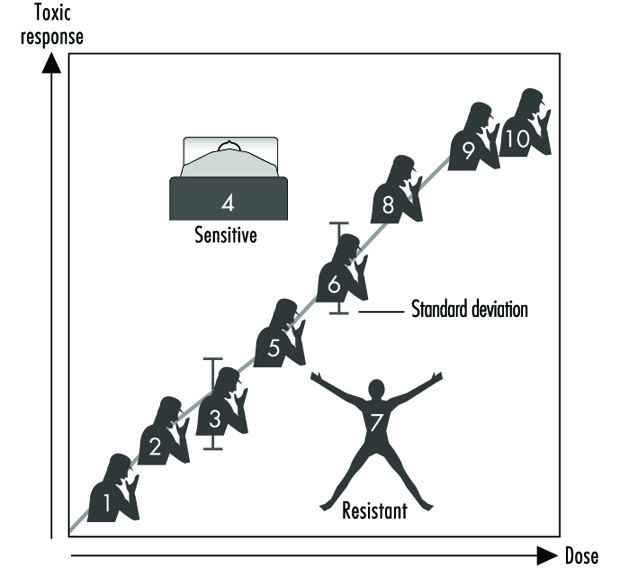
在家庭研究中使用這些異常值,許多實驗室的科學家已經開始認識到孟德爾遺傳對於給定毒性反應的重要性。 隨後,人們可以轉向分子生物學和遺傳學研究,以查明基因水平的潛在機制(基因型)負責環境引起的疾病(表型).
異生素或藥物代謝酶
身體如何應對我們接觸到的無數外源性化學物質? 人類和其他哺乳動物已經進化出高度複雜的代謝酶系統,包括十幾個不同的酶超家族。 幾乎人類接觸到的每一種化學物質都會被這些酶修飾,以促進異物從體內清除。 這些酶通常統稱為 藥物代謝酶 or 異生素代謝酶. 實際上,這兩個術語都是用詞不當。 首先,許多這些酶不僅代謝藥物,還代謝數十萬種環境和膳食化學物質。 其次,所有這些酶也有正常的身體化合物作為底物; 這些酶中沒有一種只代謝外來化學物質。
四十多年來,這些酶介導的代謝過程通常被歸類為 I 期或 II 期反應(圖 3). I 階段(“功能化”)反應通常涉及通過氧化、還原或水解對母體化學物質進行相對較小的結構修飾,以產生更易溶於水的代謝物。 通常,I 期反應為隨後的 II 期反應進一步修飾化合物提供了“把手”。 I 期反應主要由高度通用的酶超家族介導,統稱為細胞色素 P450,但也可能涉及其他酶超家族(圖 4)。

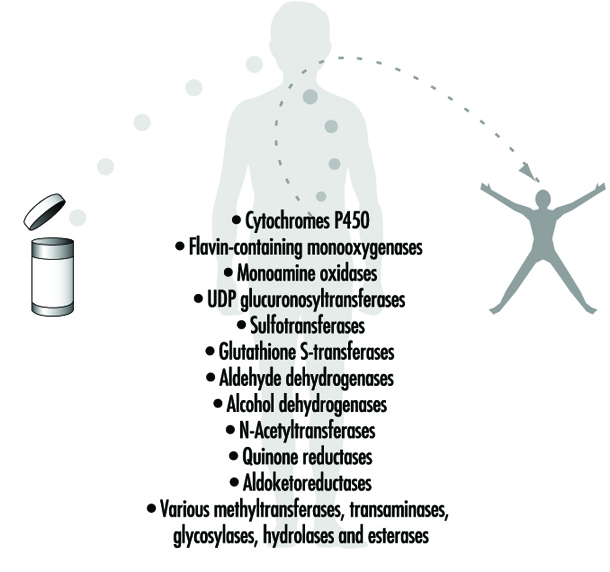
II 期反應涉及將水溶性內源性分子與化學物質(母體化學物質或 I 期代謝物)偶聯以促進排泄。 II 期反應通常稱為“偶聯”或“衍生化”反應。 催化 II 期反應的酶超家族通常根據所涉及的內源性結合部分命名:例如,N-乙酰轉移酶的乙酰化、磺基轉移酶的硫酸化、穀胱甘肽轉移酶的穀胱甘肽結合以及 UDP 葡萄醣醛酸轉移酶的葡萄醣醛酸化(圖 4) . 雖然藥物代謝的主要器官是肝臟,但某些藥物代謝酶在胃腸道、性腺、肺、腦和腎中的含量相當高,而這些酶無疑在某種程度上存在於每個活細胞中。
異生代謝酶代表雙刃劍 劍
隨著我們更多地了解導致人類健康異常的生物和化學過程,越來越明顯的是,藥物代謝酶以矛盾的方式發揮作用(圖 3)。 在大多數情況下,脂溶性化學物質會轉化為更容易排出體外的水溶性代謝物。 然而,很明顯,在許多情況下,相同的酶能夠將其他惰性化學物質轉化為高反應性分子。 然後這些中間體可以與細胞大分子如蛋白質和 DNA 相互作用。 因此,對於人類接觸的每一種化學物質,都可能存在以下競爭途徑: 代謝激活 和 排毒.
遺傳學簡述
在人類遺傳學中,每個基因(座位) 位於 23 對染色體之一。 他們倆 等位基因 (存在於一對染色體上的一個)可以相同,也可以彼此不同。 例如, B 和 b 等位基因,其中 B (棕色眼睛)占主導地位 b (藍眼睛):棕色眼睛表型的個體可以有 BB or Bb 基因型,而藍眼睛表型的個體只能具有 bb 基因型。
A 多態性 被定義為兩種或更多種穩定遺傳的表型(性狀)——源自相同的基因——在人群中得以維持,通常原因不一定明顯。 對於具有多態性的基因,基因產物必須不是發育、繁殖活力或其他關鍵生命過程所必需的。 事實上,“平衡多態性”,其中雜合子比純合子具有明顯的生存優勢(例如,抗瘧疾和鐮狀細胞血紅蛋白等位基因)是將人群中的等位基因維持在其他無法解釋的高水平的常見解釋頻率(見 岡薩雷斯和內伯特 1990 年)。
異生素代謝酶的人類多態性
四十多年來,人們已經知道各種藥物和環境化學物質代謝的遺傳差異(Kalow 1962 和 1992)。 這些差異通常被稱為 遺傳藥理學 或者,更廣泛地說, 生態遺傳多態性. 這些多態性代表在人群中以相對高的頻率出現並且通常與酶表達或功能的異常相關的變異等位基因。 從歷史上看,多態性通常是在對治療藥物產生意外反應後被發現的。 最近,重組 DNA 技術使科學家能夠確定導致其中一些多態性的基因的精確改變。 多態性現已在許多藥物代謝酶中得到表徵——包括 I 期和 II 期酶。 隨著越來越多的多態性被鑑定出來,越來越明顯的是每個個體可能擁有不同的藥物代謝酶。 這種多樣性可以描述為“代謝指紋”。 任何個體體內各種藥物代謝酶超家族的複雜相互作用最終將決定他或她對特定化學物質的特定反應(Kalow 1962 和 1992;Nebert 1988;Gonzalez 和 Nebert 1990;Nebert 和 Weber 1990)。
在細胞中表達人類異種代謝酶 文化活動
我們如何才能更好地預測人類對化學品的毒性反應? 在定義藥物代謝酶的多樣性方面取得的進展必須伴隨著關於哪些酶決定單個化學物質的代謝命運的精確知識。 從實驗室囓齒動物研究中收集的數據無疑提供了有用的信息。 然而,異生素代謝酶的種間差異顯著,因此在將數據外推到人群時必須謹慎。 為了克服這一困難,許多實驗室開發了系統,在這些系統中,可以對培養的各種細胞系進行工程改造,以產生穩定且高濃度的功能性人類酶(Gonzalez、Crespi 和 Gelboin 1991)。 已經在來自細菌、酵母、昆蟲和哺乳動物等來源的各種不同細胞系中成功生產了人類酶。
為了更準確地定義化學物質的新陳代謝, 多種酶 也已在單細胞系中成功生產(Gonzalez、Crespi 和 Gelboin 1991)。 這些細胞係為了解參與任何給定化合物和可能有毒代謝物的代謝過程的精確酶提供了寶貴的見解。 如果此信息可以與有關人體組織中酶的存在和水平的知識相結合,則這些數據應該可以提供有價值的反應預測因子。
細胞色素P450
歷史和命名
細胞色素 P450 超家族是研究最多的藥物代謝酶超家族之一,對化學物質的反應具有很大的個體差異。 細胞色素 P450 是一個方便的通用術語,用於描述在無數內源性和外源性底物的代謝中起關鍵作用的大型酶超家族。 期限 細胞色素P450 於 1962 年首次創造,用於描述未知的事物 顏料 在細胞中,當還原並與一氧化碳結合時,會在 450 nm 處產生特徵吸收峰。 自 1980 世紀 450 年代初以來,cDNA 克隆技術已經對細胞色素 P400 酶的多樣性有了顯著的了解。 迄今為止,已在動物、植物、細菌和酵母中鑑定出 450 多種不同的細胞色素 P60 基因。 據估計,任何一種哺乳動物,例如人類,可能擁有 450 種或更多不同的 P1991 基因(Nebert 和 Nelson 450)。 P1987 基因的多樣性要求開發標準化的命名系統(Nebert 等人 1993 年;Nelson 等人 1987 年)。 命名系統於 450 年首次提出並每半年更新一次,其基於 P450 蛋白之間氨基酸序列比較的發散進化。 P40 基因分為家族和亞家族:家族內的酶顯示出大於 55% 的氨基酸相似性,而同一亞家族內的酶顯示出 450% 的相似性。 PXNUMX基因以根符號命名 CYP 後面跟著一個阿拉伯數字表示 P450 家族,一個字母表示亞家族,另一個阿拉伯數字表示單個基因(Nelson et al. 1993;Nebert et al. 1991)。 因此, CYP1A1 代表家族 450 和亞家族 A 中的 P1 基因 1。
截至 1995 年 403 月,共有 XNUMX CYP 數據庫中的基因,由 59 個家族和 105 個亞家族組成。 其中包括 15 個低等真核生物科、19 個植物科和 15 個細菌科。 450 個人類 P26 基因家族包含 22 個亞家族,其中 XNUMX 個已被定位到整個基因組的大部分染色體位置。 一些序列在許多物種中顯然是同源的——例如,只有一個 CYP17 (類固醇 17α-羥化酶)基因已在迄今為止檢查的所有脊椎動物中發現; 一個亞家族中的其他序列高度重複,使得不可能識別直系同源對(例如, CYP2C 亞科)。 有趣的是,人類和酵母在 CYP51 家庭。 為尋求 P450 超家族更多信息的讀者提供了大量綜合評論(Nelson 等人 1993 年;Nebert 等人 1991 年;Nebert 和 McKinnon 1994 年;Guengerich 1993 年;Gonzalez 1992 年)。
P450 命名系統的成功導致為 UDP 葡萄醣醛酸轉移酶(Burchell 等人,1991 年)和含黃素的單加氧酶(Lawton 等人,1994 年)開發了類似的術語系統。 其他幾個藥物代謝酶超家族(例如,磺基轉移酶、環氧化物水解酶和醛脫氫酶)也正在開發基於發散進化的類似命名系統。
最近,我們將哺乳動物 P450 基因超家族分為三組(Nebert 和 McKinnon 1994)——主要涉及外來化學代謝的組、參與各種類固醇激素合成的組以及參與其他重要內源性功能的組。 外源代謝 P450 酶對毒性預測最重要。
異生素代謝 P450 酶
參與外來化合物和藥物代謝的 P450 酶幾乎總是在家族中發現 CYP1、CYP2、CYP3 和 CYP4. 這些 P450 酶催化多種代謝反應,單個 P450 通常能夠代謝許多不同的化合物。 此外,多種 P450 酶可能會在不同位點代謝單一化合物。 此外,一種化合物可能在同一個位點被多個 P450 代謝,儘管速率不同。
藥物代謝 P450 酶的一個最重要的特性是這些基因中的許多都可以被用作其底物的物質誘導。 另一方面,其他 P450 基因是由非底物誘導的。 這種酶誘導現像是許多具有治療重要性的藥物相互作用的基礎。
儘管存在於許多組織中,但這些特殊的 P450 酶在肝臟(藥物代謝的主要部位)中含量相對較高。 一些代謝外源性物質的 P450 酶對某些內源性底物(例如花生四烯酸)表現出活性。 然而,人們普遍認為,這些代謝外源性物質的 P450 酶中的大多數並不發揮重要的生理作用——儘管這尚未通過實驗確定。 通過基因打靶方法在小鼠中選擇性純合子破壞或“敲除”單個異源代謝 P450 基因可能會很快提供關於異源代謝 P450 的生理作用的明確信息(用於回顧基因打靶,參見 Capecchi 1994)。
與編碼主要參與生理過程的酶的 P450 家族相比,編碼異生物質代謝 P450 酶的家族顯示出顯著的物種特異性,並且每個亞家族通常包含許多活性基因(Nelson 等人,1993 年;Nebert 等人,1991 年)。 鑑於明顯缺乏生理底物,家族中的 P450 酶可能 CYP1、CYP2、CYP3 和 CYP4 在過去數億年中出現的化學物質已經進化為一種對環境和飲食中遇到的外來化學物質進行解毒的方法。 顯然,異生素代謝 P450 的進化發生的時間遠遠早於人類現在接觸的大多數合成化學物質的合成。 這四個基因家族中的基因可能由於動物在過去 1.2 億年中接觸植物代謝物而進化和分化——這一過程被描述為“動植物戰爭”(Gonzalez 和 Nebert,1990 年)。 動植物戰爭是一種現象,其中植物開發出新的化學物質(植物抗毒素)作為防禦機制以防止被動物攝入,而動物反過來又通過開發新的 P450 基因來適應多樣化的底物。 最近描述的涉及 P450 有毒底物解毒的植物-昆蟲和植物-真菌化學戰的例子進一步推動了該提議(Nebert 1994)。
以下是對幾種人類異源代謝 P450 酶多態性的簡要介紹,其中毒性反應的遺傳決定因素被認為具有重要意義。 直到最近,P450 多態性通常是由患者對所用治療劑的反應出乎意料的差異所暗示的。 確實有幾個 P450 多態性是根據首次鑑定該多態性的藥物命名的。 最近,研究工作的重點是鑑定參與化學物質代謝的精確 P450 酶,觀察到這些酶存在差異,並精確表徵所涉及的 P450 基因。 如前所述,P450 酶對模型化學物質的可測量活性可稱為表型。 每個個體的 P450 基因的等位基因差異稱為 P450 基因型。 隨著越來越多的審查應用於 P450 基因的分析,先前記錄的表型變異的精確分子基礎變得更加清晰。
CYP1A亞家族
CYP1A 亞家族包含人類和所有其他哺乳動物中的兩種酶:這些酶在標準 P1 命名法下被命名為 CYP1A1 和 CYP2A450。 這些酶很受關注,因為它們參與許多致癌物的代謝活化,並且還被包括二噁英在內的幾種具有毒理學關注的化合物誘導。 例如,CYP1A1 代謝激活香煙煙霧中發現的許多化合物。 CYP1A2 代謝激活化學染料工業中發現的許多與膀胱癌相關的芳基胺。 CYP1A2 還通過代謝激活 4-(甲基亞硝胺)-1-(3-吡啶基)-1-丁酮 (NNK),一種源自煙草的亞硝胺。 由於菸霧中存在的多環碳氫化合物的誘導,CYP1A1 和 CYP1A2 在吸煙者肺部的含量也較高。 因此,CYP1A1 和 CYP1A2 活性水平被認為是個體對許多潛在有毒化學品反應的重要決定因素。
毒理學興趣 CYP1A 1973 年的一份報告將吸煙者的 CYP1A1 誘導水平與個人對肺癌的易感性相關聯(Kellermann、Shaw 和 Luyten-Kellermann 1973)。 CYP1A1 和 CYP1A2 誘導的分子基礎一直是眾多實驗室關注的焦點。 誘導過程由一種稱為 Ah 受體的蛋白質介導,二噁英和結構相關的化學物質與其結合。 名字 Ah 源自 a瑞爾 h許多 CYP1A 誘導劑的碳氫化合物性質。 有趣的是,小鼠品系之間編碼 Ah 受體的基因的差異導致化學反應和毒性的顯著差異。 Ah 受體基因的多態性似乎也發生在人類身上:大約十分之一的人群表現出 CYP1A1 的高誘導性,並且可能比其他十分之九的人群更容易患某些化學誘發的癌症。 Ah 受體在酶控制中的作用 CYP1A 亞家族及其作為人類對化學暴露反應的決定因素的作用,一直是最近幾篇評論的主題(Nebert、Petersen 和 Puga 1991;Nebert、Puga 和 Vasiliou 1993)。
是否有其他多態性可能控制細胞中 CYP1A 蛋白的水平? 中的多態性 CYP1A1 基因也已被確定,這似乎會影響日本吸煙者的肺癌風險,儘管這種相同的多態性似乎不會影響其他種族群體的風險(Nebert 和 McKinnon 1994)。
CYP2C19
多年來,人們對抗驚厥藥物 (S)-美芬妥英的代謝率存在差異已有詳細記載 (Guengerich 1989)。 2% 至 5% 的白種人和多達 25% 的亞洲人缺乏這種活性,並且可能面臨更大的藥物毒性風險。 人們早就知道這種酶缺陷涉及人類的一個成員 CYP2C 亞家族,但這種缺陷的精確分子基礎一直是相當大的爭議主題。 造成這種困難的主要原因是人類的六個或更多基因 CYP2C 亞科。 然而,最近證明,在 CYP2C19 基因是造成這種缺陷的主要原因 (Goldstein and de Morais 1994)。 還開發了一種基於聚合酶鍊式反應 (PCR) 的簡單 DNA 測試,以快速識別人群中的這種突變(Goldstein 和 de Morais 1994)。
CYP2D6
也許 P450 基因中最廣泛表徵的變異是涉及 CYP2D6 基因。 已經描述了十多個影響該基因的突變、重排和缺失的例子 (Meyer 1994)。 20 年前,患者對抗高血壓藥物異喹啉的反應存在臨床差異,首次提出了這種多態性。 的改變 CYP2D6 因此,引起酶活性改變的基因統稱為 異喹多態性.
在基於 DNA 的研究出現之前,根據尿液樣本中的代謝物濃度,個體被歸類為異喹啉的弱代謝者或強代謝者(PM、EM)。 現在很清楚 CYP2D6 基因可能導致個體不僅表現出不良或廣泛的異喹啉代謝,而且表現出超快的代謝。 大多數改動在 CYP2D6 基因與酶功能的部分或全部缺陷有關; 然而,最近描述了兩個家庭中的個體擁有多個功能副本 CYP2D6 基因,引起 CYP2D6 底物的超快速代謝(Meyer 1994)。 這一非凡的觀察為以前在人口研究中觀察到的廣泛的 CYP2D6 活性提供了新的見解。 CYP2D6 功能的改變具有特別重要的意義,因為該酶可代謝 30 多種常用處方藥。 因此,個體的 CYP2D6 功能是對給藥治療的治療反應和毒性反應的主要決定因素。 事實上,最近有人認為,考慮患者的 CYP2D6 狀態對於安全使用精神科和心血管藥物是必要的。
的作用 CYP2D6 多態性作為個體對人類疾病如肺癌和帕金森病的易感性的決定因素也一直是深入研究的主題(Nebert 和 McKinnon 1994 年;Meyer 1994 年)。 雖然鑑於所用研究方案的多樣性,很難得出結論,但大多數研究似乎表明異喹啉的廣泛代謝者(EM 表型)與肺癌之間存在關聯。 這種關聯的原因目前尚不清楚。 然而,CYP2D6 酶已顯示可代謝 NNK,一種源自煙草的亞硝胺。
隨著基於 DNA 的檢測方法的改進——能夠更準確地評估 CYP2D6 狀態——預計 CYP2D6 與疾病風險的確切關係將得到闡明。 強代謝者可能與肺癌易感性有關,而弱代謝者(PM 表型)似乎與原因不明的帕金森病有關。 儘管這些研究也難以比較,但似乎代謝 CYP2D6 底物(例如,異喹啉)能力降低的 PM 個體患帕金森病的風險增加 2 至 2.5 倍。
CYP2E1
CYP2E1 基因編碼的酶可以代謝許多化學物質,包括藥物和許多低分子量致癌物。 這種酶也很受關注,因為它可被酒精高度誘導,並且可能在氯仿、氯乙烯和四氯化碳等化學物質引起的肝損傷中發揮作用。 這種酶主要存在於肝臟中,酶的水平因人而異。 仔細檢查 CYP2E1 基因導致了幾種多態性的鑑定(Nebert 和 McKinnon 1994)。 據報導,某些結構變異的存在之間存在關係 CYP2E1 一些研究中的基因和明顯降低的肺癌風險; 然而,種族間存在明顯差異,需要澄清這種可能的關係。
CYP3A亞家族
在人類中,四種酶已被確定為 CYP3A 亞科是因為它們在氨基酸序列上的相似性。 CYP3A 酶代謝許多常用處方藥,例如紅黴素和環孢菌素。 致癌食品污染物黃曲霉毒素 B1 也是 CYP3A 底物。 人類的一員 CYP3A 亞科,指定 CYP3A4, 是人類肝臟中的主要 P450,也存在於胃腸道中。 與許多其他 P450 酶一樣,CYP3A4 的水平在個體之間差異很大。 第二種酶,命名為 CYP3A5,僅存在於大約 25% 的肝臟中; 這一發現的遺傳基礎尚未闡明。 CYP3A4 或 CYP3A5 變異性作為毒性反應遺傳決定因素的重要性尚未確定(Nebert 和 McKinnon 1994)。
非 P450 多態性
許多多態性也存在於其他異源代謝酶超家族中(例如,穀胱甘肽轉移酶、UDP 葡萄醣醛酸轉移酶、對氧磷酶、脫氫酶、N-乙酰轉移酶和含黃素的單加氧酶)。 由於任何 P450 生成的中間體的最終毒性取決於隨後的 II 期解毒反應的效率,因此多種酶多態性的綜合作用對於確定對化學誘發疾病的易感性很重要。 因此,I 期和 II 期反應之間的代謝平衡(圖 3)可能是化學誘發的人類疾病和毒性反應的遺傳決定因素的主要因素。
GSTM1基因多態性
II 期酶多態性的一個充分研究的例子是涉及穀胱甘肽 S-轉移酶超家族的成員,指定為 GST mu 或 GSTM1。 這種特殊的酶具有相當大的毒理學意義,因為它似乎參與了 CYP1A1 酶對香煙煙霧中化學物質產生的有毒代謝物的後續解毒。 這種穀胱甘肽轉移酶基因的多態性涉及在多達一半的所有研究的白種人中完全缺乏功能性酶。 這種 II 相酶的缺乏似乎與肺癌易感性增加有關。 通過基於兩種變體對個體進行分組 CYP1A1 基因和功能缺失或存在 GSTM1 基因,已證明患吸煙誘發肺癌的風險存在顯著差異(Kawajiri、Watanabe 和 Hayashi 1994)。 特別是,個人展示了一種罕見的 CYP1A1 基因改變,結合缺乏 GSTM1 基因,當暴露於相對低水平的香煙煙霧時,患肺癌的風險更高(高達九倍)。 有趣的是,變異基因的重要性似乎存在種族間差異,這需要進一步研究以闡明此類改變在疾病易感性中的確切作用(Kalow 1962;Nebert 和 McKinnon 1994;Kawajiri、Watanabe 和 Hayashi 1994)。
兩種或多種多態性對毒性的協同作用 響應
對環境因素的毒性反應可能會因同一個體中兩種藥物遺傳學缺陷的組合而大大加劇,例如,N-乙酰轉移酶 (NAT2) 多態性和葡萄糖-6-磷酸脫氫酶 (G6PD) 多態性的聯合作用.
職業性接觸芳基胺是患膀胱癌的嚴重風險。 自 1954 年 Cartwright 的優雅研究以來,很明顯 N-乙酰基狀態是偶氮染料誘發膀胱癌的決定因素。 慢乙酰化表型與膀胱癌的發生以及這種癌症在膀胱壁的侵襲程度之間存在高度顯著的相關性。 相反,快速乙酰表型與結直腸癌的發病率之間存在顯著關聯。 N-乙酰轉移酶(NAT1、NAT2) 基因已被克隆和測序,基於 DNA 的分析現在能夠檢測出十多種等位基因變異,這些變異是慢乙酰化表型的原因。 這 NAT2 基因是多態性的,並且造成對環境化學物質的毒性反應的大部分變異性(Weber 1987;Grant 1993)。
6-磷酸葡萄糖脫氫酶 (G6PD) 是一種對 NADPH 的生成和維持至關重要的酶。 由於紅細胞中缺乏正常水平的還原型穀胱甘肽 (GSH),G6PD 活性低或不存在可導致嚴重的藥物或異物誘導的溶血。 G6PD 缺乏症影響著全世界至少 300 億人。 超過 10% 的非裔美國男性表現出不太嚴重的表型,而某些撒丁島社區表現出更嚴重的“地中海型”,頻率高達每三個人中就有一個。 這 G6PD 基因已被克隆並定位於 X 染色體,大量不同的點突變導致了 G6PD 缺陷個體中出現的大量表型異質性 (Beutler 1992)。
Thiozalsulphone 是一種芳胺磺胺藥物,被發現會在接受治療的人群中引起溶血性貧血的雙峰分佈。 當用某些藥物治療時,同時具有 G6PD 缺乏症和慢乙酰化表型的個體比僅具有 G6PD 缺乏症或僅具有慢乙酰化表型的個體受到的影響更大。 G6PD 缺陷型慢速乙酰化器對硫唑砜誘導的溶血的敏感性至少是正常 G40PD 快速乙酰化器的 6 倍。
遺傳多態性對暴露評估的影響
暴露評估和生物監測(圖 1)也需要每個人的基因構成信息。 鑑於相同的危險化學品暴露,血紅蛋白加合物(或其他生物標誌物)的水平可能在個體之間相差兩到三個數量級,這取決於每個人的代謝指紋。
在德國的化工廠工人中研究了相同的組合遺傳藥理學(表 1)。 與其他可能的組合藥物遺傳學表型相比,暴露於苯胺和乙酰苯胺的工人中的血紅蛋白加合物在 G6PD 缺陷型慢乙酰化器中是迄今為止最高的。 這項研究對暴露評估具有重要意義。 這些數據表明,儘管兩個人可能在工作場所暴露於相同環境水平的危險化學品,但暴露量(通過血紅蛋白加合物等生物標誌物)可能估計要低兩個或更多數量級,因為與個體潛在的遺傳易感性有關。 同樣,由此產生的不利健康影響的風險可能相差兩個或更多數量級。
表 1:接觸苯胺和乙酰苯胺的工人的血紅蛋白加合物
| 乙酰化狀態 | G6PD缺乏症 | |||
| 快 | 放慢 | 沒有 | 是 | 血紅蛋白加合物 |
| + | + | 2 | ||
| + | + | 30 | ||
| + | + | 20 | ||
| + | + | 100 | ||
資料來源:改編自 Lewalter 和 Korallus 1985。
結合和代謝的遺傳差異
應該強調的是,此處針對新陳代謝提出的相同案例也適用於結合。 環境因素結合的遺傳差異將極大地影響毒性反應。 例如,鼠標的差異 清潔劑 基因可以深刻地影響個體對鎘誘導的睾丸壞死的敏感性(Taylor、Heiniger 和 Meier 1973)。 Ah 受體結合親和力的差異可能會影響二噁英誘導的毒性和癌症(Nebert、Petersen 和 Puga 1991 年;Nebert、Puga 和 Vasiliou 1993 年)。
圖 5 總結了代謝和結合在毒性和癌症中的作用。 有毒物質,當它們存在於環境中或在代謝或結合後,通過基因毒性途徑(其中發生 DNA 損傷)或非基因毒性途徑(其中不需要發生 DNA 損傷和誘變)引起它們的作用。 有趣的是,最近很明顯,“經典”DNA 損傷劑可以通過還原型穀胱甘肽 (GSH) 依賴性非遺傳毒性信號轉導通路發揮作用,該通路在沒有 DNA 的情況下在細胞核外的細胞表面或附近啟動(Devary 等人,1993 年)。 然而,代謝和結合的遺傳差異仍然是控制不同個體毒性反應的主要決定因素。
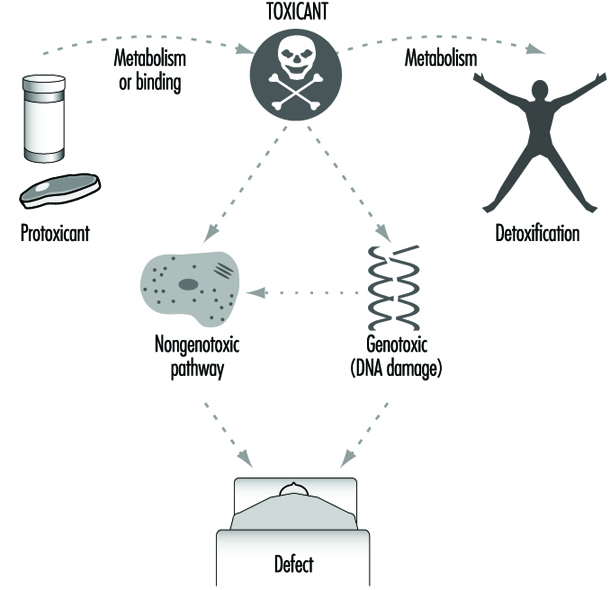
藥物代謝酶在細胞功能中的作用
藥物代謝酶功能的遺傳變異對於確定個體對化學品的反應至關重要。 這些酶對於確定接觸外來化學物質後的命運和時間過程至關重要。
如圖 5 所示,藥物代謝酶在個體對化學暴露的易感性中的重要性實際上可能提出了一個比從這個簡單的異生物質代謝討論中顯而易見的問題複雜得多的問題。 換句話說,在過去的二十年中,基因毒性機制(DNA 加合物和蛋白質加合物的測量)得到了極大的強調。 然而,如果非基因毒性機制在引起毒性反應方面至少與基因毒性機制一樣重要呢?
如前所述,許多參與異生素代謝的藥物代謝酶的生理作用尚未準確定義。 Nebert (1994) 提出,由於藥物代謝酶在這個星球上存在超過 3.5 億年,它們最初(現在仍然主要)負責調節許多對轉錄激活很重要的非肽配體的細胞水平影響生長、分化、凋亡、穩態和神經內分泌功能的基因。 此外,大多數(如果不是全部的話)環境因素的毒性是通過 激動劑 or 對手 對這些信號轉導通路的作用 (Nebert 1994)。 基於這一假設,藥物代謝酶的遺傳變異性可能對細胞內的許多關鍵生化過程產生相當顯著的影響,從而導致毒性反應的重要差異。 這種情況確實有可能也是使用常用處方藥的患者遇到的許多特殊不良反應的基礎。
結論
在過去的十年裡,我們對藥物、食品和環境污染物中化學物質的不同反應的遺傳基礎的理解取得了顯著進步。 藥物代謝酶對人類對化學品的反應方式有著深遠的影響。 隨著我們對藥物代謝酶多樣性的認識不斷發展,我們越來越能夠改進許多藥物和環境化學品的毒性風險評估。 這可能在 CYP2D6 細胞色素 P450 酶的情況下得到了最清楚的說明。 使用相對簡單的基於 DNA 的測試,可以預測主要由這種酶代謝的任何藥物的可能反應; 這一預測將確保更安全地使用有價值但可能有毒的藥物。
未來無疑會看到涉及藥物代謝酶的進一步多態性(表型)鑑定的爆炸式增長。 這些信息將伴隨改進的、微創的基於 DNA 的測試來識別人群中的基因型。
這些研究在評估化學品在目前未知來源的許多環境疾病中的作用方面應該特別有用。 綜合考慮多種藥物代謝酶多態性(如表 1),也可能代表一個特別豐富的研究領域。 這些研究將闡明化學物質在致癌中的作用。 總的來說,這些信息應該有助於製定越來越個性化的建議,避免可能引起個人關注的化學品。 這是預防性毒理學領域。 這樣的建議無疑將極大地幫助所有人應對我們所面臨的不斷增加的化學負擔。