幾乎所有的醫學都致力於預防細胞死亡,如心肌梗塞、中風、外傷和休克等疾病,或引起細胞死亡,如傳染病和癌症。 因此,必須了解所涉及的性質和機制。 細胞死亡被歸類為“意外”,即由有毒物質、局部缺血等引起,或“程序性”,發生在胚胎髮育過程中,包括手指的形成和蝌蚪尾巴的吸收。
因此,細胞損傷和細胞死亡在生理學和病理生理學中都很重要。 生理細胞死亡在胚胎髮生和胚胎髮育過程中極為重要。 對發育過程中細胞死亡的研究導致了有關分子遺傳學的重要和新信息,特別是通過對無脊椎動物發育的研究。 在這些動物中,已經仔細研究了注定要經歷細胞死亡的細胞的精確位置和重要性,並且通過使用經典的誘變技術,現在已經確定了幾個相關基因。 在成人器官中,細胞死亡和細胞增殖之間的平衡控制著器官的大小。 在一些器官中,例如皮膚和腸道,細胞不斷更新。 例如,在皮膚中,細胞在到達表面時會分化,並最終隨著角質化的進行以及交聯包膜的形成而經歷終末分化和細胞死亡。
許多類別的有毒化學品都能夠誘導急性細胞損傷,然後導致死亡。 這些包括缺氧和局部缺血及其化學類似物,例如氰化鉀; 化學致癌物,形成與核酸中蛋白質共價結合的親電體; 氧化劑化學品,導致自由基形成和氧化損傷; 激活補體; 和多種鈣離子載體。 細胞死亡也是化學致癌作用的重要組成部分; 許多完全的化學致癌物在致癌劑量下會產生急性壞死和炎症,然後是再生和癌前病變。
定義
細胞損傷
細胞損傷被定義為擾亂細胞正常穩態的事件或刺激,例如有毒化學物質,從而導致許多事件的發生(圖 1)。 所示致死性損傷的主要目標是 ATP 合成的抑制、質膜完整性的破壞或必需生長因子的撤回。
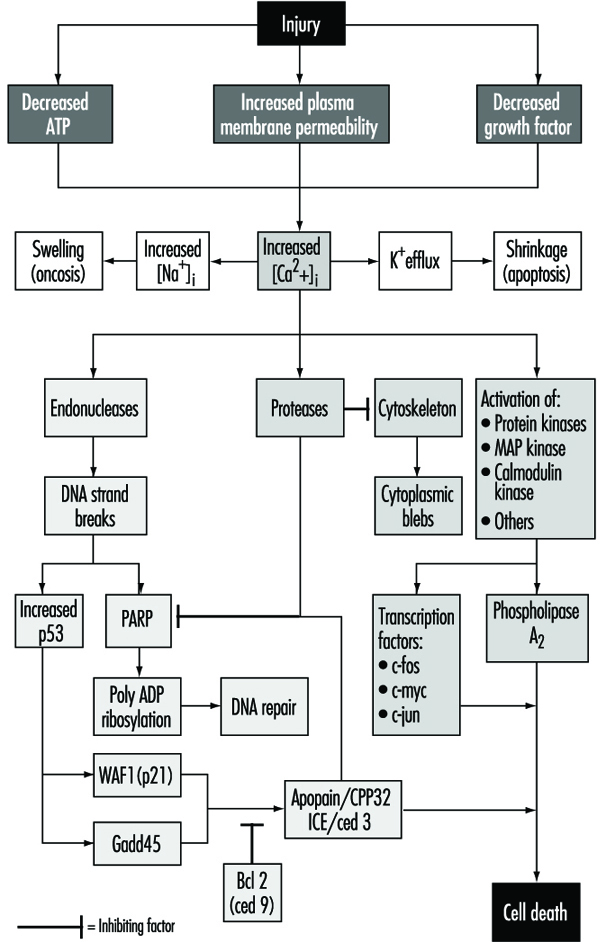
致命傷害會導致細胞在一段不同的時間後死亡,這取決於溫度、細胞類型和刺激; 或者它們可能是亞致死性或慢性的——也就是說,損傷會導致體內平衡狀態改變,雖然不正常,但不會導致細胞死亡(Trump 和 Arstila 1971;Trump 和 Berezesky 1992;Trump 和 Berezesky 1995;Trump、Berezesky 和Osornio-Vargas 1981)。 在致命傷害的情況下,在細胞死亡之前有一個階段
在此期間,細胞會恢復; 然而,在特定的時間點(“不歸路點”或細胞死亡點)之後,損傷的消除並不會導致恢復,而是細胞會發生降解和水解,最終與細胞達到物理化學平衡環境。 這是稱為壞死的階段。 在致死前階段,會發生幾種主要類型的變化,具體取決於細胞和損傷類型。 這些被稱為細胞凋亡和腫瘤。
細胞凋亡
Apoptosis 源自希臘語 載脂蛋白,意思是遠離,並且 下垂,跌倒的意思。 期限 遠離 源於這樣一個事實,即在這種類型的致死前變化期間,細胞會收縮並在外圍經歷明顯的起泡。 然後氣泡分離並漂浮。 細胞凋亡發生在各種類型的中毒性損傷後的各種細胞類型中(Wyllie、Kerr 和 Currie 1980)。 它在淋巴細胞中尤為突出,它是淋巴細胞克隆周轉的主要機制。 由此產生的片段導致在淋巴結中的巨噬細胞內看到嗜鹼性體。 在其他器官中,細胞凋亡通常發生在單個細胞中,這些細胞在死亡前後被鄰近的實質細胞或巨噬細胞的片段吞噬作用迅速清除。 在單細胞中發生的細胞凋亡以及隨後的吞噬作用通常不會導致炎症。 在死亡之前,凋亡細胞顯示出非常緻密的胞質溶膠和正常或濃縮的線粒體。 內質網 (ER) 正常或僅輕微擴張。 核染色質沿著核膜和核仁周圍明顯聚集。 核輪廓也不規則,發生核碎裂。 染色質濃縮與 DNA 片段化有關,在許多情況下,DNA 片段化發生在核小體之間,在電泳中呈現出特徵性的階梯狀外觀。
在細胞凋亡中,[Ca2+]i 可能刺激 K+ 流出導致細胞收縮,這可能需要 ATP。 因此,完全抑制 ATP 合成的損傷更有可能導致細胞凋亡。 [Ca 的持續增加2+]i 具有許多有害作用,包括激活蛋白酶、核酸內切酶和磷脂酶。 核酸內切酶激活導致單鍊和雙鏈 DNA 斷裂,進而刺激 p53 和多聚 ADP 核糖基化以及 DNA 修復所必需的核蛋白水平升高。 蛋白酶的激活會改變許多底物,包括肌動蛋白和導致水泡形成的相關蛋白質。 另一個重要的底物是聚(ADP-核糖)聚合酶 (PARP),它會抑制 DNA 修復。 增加 [Ca2+]i 也與許多蛋白激酶的激活有關,例如 MAP 激酶、鈣調蛋白激酶等。 此類激酶參與啟動立即早期基因轉錄的轉錄因子的激活,例如,c-fos、c-jun 和 c-myc,以及磷脂酶 A 的激活2 這導致質膜和細胞內膜(例如線粒體內膜)的透化。
腫瘤病
Oncosis,源自希臘詞 昂科斯腫脹之所以如此命名,是因為在這種類型的致死前變化中,細胞在受傷後幾乎立即開始腫脹(Majno 和 Joris 1995)。 膨脹的原因是細胞內水中陽離子的增加。 負責的主要陽離子是鈉,它通常被調節以維持細胞體積。 然而,在沒有 ATP 的情況下或如果質膜的 Na-ATPase 受到抑制,由於細胞內蛋白質和水中的鈉繼續增加,體積控制失去。 因此,在腫瘤病的早期事件中,[Na+]i 這導致細胞腫脹和增加 [Ca2+]i 由細胞外空間流入或細胞內儲存釋放引起。 這導致胞質溶膠腫脹、內質網和高爾基體腫脹,以及在細胞表面周圍形成水泡。 線粒體最初經歷濃縮,但後來由於線粒體內膜受損,它們也表現出高幅度的腫脹。 在這種致死前的變化中,染色質經歷濃縮並最終降解; 然而,沒有看到細胞凋亡的特徵階梯模式。
壞疽
壞死是指細胞死亡後發生的一系列變化,當細胞轉化為通常通過炎症反應去除的碎片時。 可以區分兩種類型:膨脹性壞死和凋亡性壞死。 腫瘤性壞死通常發生在大面積區域,例如,心肌梗塞或化學中毒後的局部器官,例如施用 HgCl 後的腎近端小管2. 涉及器官的廣泛區域,壞死細胞迅速引發炎症反應,首先是急性的,然後是慢性的。 在生物體存活的情況下,在許多器官壞死之後會清除死細胞並再生,例如,在化學毒性後的肝臟或腎臟中。 相反,凋亡性壞死通常發生在單個細胞的基礎上,壞死碎片在巨噬細胞的吞噬細胞或鄰近的實質細胞內形成。 壞死細胞的最早特徵包括質膜連續性中斷和絮狀密度的出現,代表線粒體基質內的變性蛋白質。 在最初不干擾線粒體鈣積累的某些形式的損傷中,可以在線粒體中看到磷酸鈣沉積物。 其他膜系統也有類似的碎片化,例如內質網、溶酶體和高爾基體。 最終,由於溶酶體水解酶的攻擊,核染色質發生裂解。 細胞死亡後,溶酶體水解酶在與組織蛋白酶、核糖酶和脂肪酶一起清除碎片中發揮重要作用,因為它們具有最佳酸性 pH 值,並且可以在壞死細胞的低 pH 值下存活,而其他細胞酶則變性和失活。
機制
初始刺激
在致命傷害的情況下,最常見的導致細胞死亡的傷害的初始相互作用是乾擾能量代謝,例如缺氧、局部缺血或呼吸抑制,以及糖酵解,例如氰化鉀、一氧化碳、碘乙酸鹽和很快。 如上所述,抑制能量代謝的高劑量化合物通常會導致腫瘤。 導致急性細胞死亡的另一種常見初始損傷類型是質膜功能的改變(Trump 和 Arstila 1971;Trump、Berezesky 和 Osornio-Vargas 1981)。 這可能是直接損傷和透化作用,如外傷或補體 C5b-C9 複合物激活、細胞膜機械損傷或鈉鉀 (Na+-K+) 與哇巴因等苷類一起泵送。 鈣離子載體,如離子黴素或 A23187,可快速攜帶 [Ca2+] 向下梯度進入細胞,也會造成急性致死性損傷。 在某些情況下,致死前變化的模式是細胞凋亡; 在其他情況下,它是腫瘤。
信號通路
對於多種類型的損傷,線粒體呼吸和氧化磷酸化會迅速受到影響。 在某些細胞中,這會刺激能夠維持 ATP 的無氧糖酵解,但在許多損傷中,這會受到抑制。 缺乏 ATP 會導致無法激活許多重要的穩態過程,特別是細胞內離子穩態的控制(Trump 和 Berezesky 1992;Trump、Berezesky 和 Osornio-Vargas 1981)。 這導致 [Ca2+]i, 並增加 [Na+] 和 [Cl-]導致細胞腫脹。 [Ca 增加2+]i 導致下面討論的許多其他信號機制的激活,包括一系列激酶,這可能導致立即早期基因轉錄增加。 增加 [Ca2+]i 還改變細胞骨架功能,部分導致氣泡形成和核酸內切酶、蛋白酶和磷脂酶的激活。 這些似乎觸發了上面討論的許多重要影響,例如通過蛋白酶和脂肪酶激活引起的膜損傷,核酸內切酶激活導致的 DNA 直接降解,以及作為轉錄因子的 MAP 激酶和鈣調蛋白激酶等激酶的激活。
通過對無脊椎動物發育的廣泛研究 秀麗隱桿線蟲 和 果蠅以及人類和動物細胞,已經確定了一系列促死亡基因。 已發現其中一些無脊椎動物基因具有哺乳動物對應物。 例如,ced-3 基因,它對細胞程序性死亡至關重要 秀麗隱桿線蟲, 具有蛋白酶活性並與哺乳動物白細胞介素轉化酶 (ICE) 具有很強的同源性。 最近已鑑定出一種密切相關的基因,稱為 apopain 或 prICE,具有更緊密的同源性 (Nicholson et al. 1995)。 在 果蠅,收割者基因似乎參與了導致程序性細胞死亡的信號。 其他促死亡基因包括 Fas 膜蛋白和重要的腫瘤抑制基因 p53,後者被廣泛保存。 p53 在 DNA 損傷後在蛋白質水平上被誘導,當磷酸化作為其他基因(如 gadd45 和 waf-1)的轉錄因子時,這些基因參與細胞死亡信號傳導。 其他直接早期基因如 c-fos、c-jun 和 c-myc 似乎也參與某些系統。
同時,還有一些抗死亡基因似乎可以抵消促死亡基因。 其中第一個被識別的是 ced-9,來自 秀麗隱桿線蟲,與人類的 bcl-2 同源。 這些基因以一種未知的方式起作用,以防止細胞被遺傳或化學毒素殺死。 最近的一些證據表明 bcl-2 可以作為一種抗氧化劑。 目前,人們正在努力加深對相關基因的了解,並根據情況開發激活或抑制這些基因的方法。