Il est reconnu depuis longtemps que la réaction de chaque personne aux produits chimiques environnementaux est différente. L'explosion récente de la biologie moléculaire et de la génétique a permis de mieux comprendre les bases moléculaires d'une telle variabilité. Les principaux déterminants de la réponse individuelle aux produits chimiques comprennent des différences importantes entre plus d'une douzaine de superfamilles d'enzymes, appelées collectivement xénobiotique- (étranger au corps) ou métabolisant les médicaments enzymes. Bien que le rôle de ces enzymes ait été classiquement considéré comme une détoxification, ces mêmes enzymes convertissent également un certain nombre de composés inertes en intermédiaires hautement toxiques. Récemment, de nombreuses différences aussi bien subtiles que grossières dans les gènes codant pour ces enzymes ont été identifiées, lesquelles se sont avérées entraîner des variations marquées de l'activité enzymatique. Il est maintenant clair que chaque individu possède un complément distinct d'activités enzymatiques métabolisant les xénobiotiques ; cette diversité pourrait être considérée comme une « empreinte métabolique ». C'est l'interaction complexe de ces nombreuses superfamilles d'enzymes différentes qui détermine en fin de compte non seulement le devenir et le potentiel de toxicité d'un produit chimique chez un individu donné, mais également l'évaluation de l'exposition. Dans cet article, nous avons choisi d'utiliser la superfamille des enzymes du cytochrome P450 pour illustrer les progrès remarquables réalisés dans la compréhension de la réponse individuelle aux produits chimiques. Le développement de tests basés sur l'ADN relativement simples conçus pour identifier des altérations génétiques spécifiques dans ces enzymes fournit désormais des prédictions plus précises de la réponse individuelle à l'exposition chimique. Nous espérons que le résultat sera une toxicologie préventive. En d'autres termes, chaque individu pourrait en apprendre davantage sur les produits chimiques auxquels il est particulièrement sensible, évitant ainsi une toxicité ou un cancer auparavant imprévisible.
Bien que cela ne soit généralement pas apprécié, les êtres humains sont exposés quotidiennement à un barrage d'innombrables produits chimiques divers. Bon nombre de ces produits chimiques sont hautement toxiques et proviennent d'une grande variété de sources environnementales et alimentaires. La relation entre ces expositions et la santé humaine a été, et continue d'être, un objectif majeur des efforts de recherche biomédicale dans le monde entier.
Quels sont quelques exemples de ce bombardement chimique ? Plus de 400 substances chimiques du vin rouge ont été isolées et caractérisées. On estime qu'au moins 1,000 75,000 produits chimiques sont produits par une cigarette allumée. Il existe d'innombrables produits chimiques dans les cosmétiques et les savons parfumés. L'agriculture est une autre source majeure d'exposition aux produits chimiques : rien qu'aux États-Unis, les terres agricoles reçoivent chaque année plus de XNUMX XNUMX produits chimiques sous forme de pesticides, d'herbicides et d'agents fertilisants ; après absorption par les plantes et les animaux au pâturage, ainsi que par les poissons dans les cours d'eau à proximité, les humains (à la fin de la chaîne alimentaire) ingèrent ces produits chimiques. Deux autres sources de fortes concentrations de produits chimiques absorbés par le corps comprennent (a) les médicaments pris de manière chronique et (b) l'exposition à des substances dangereuses sur le lieu de travail au cours d'une vie d'emploi.
Il est désormais bien établi que l'exposition aux produits chimiques peut avoir des effets néfastes sur de nombreux aspects de la santé humaine, provoquant des maladies chroniques et le développement de nombreux cancers. Au cours de la dernière décennie, la base moléculaire de bon nombre de ces relations a commencé à être dévoilée. De plus, la prise de conscience a émergé que les humains diffèrent nettement dans leur susceptibilité aux effets nocifs de l'exposition chimique.
Les efforts actuels pour prédire la réponse humaine à l'exposition chimique combinent deux approches fondamentales (figure 1) : surveiller l'étendue de l'exposition humaine à l'aide de marqueurs biologiques (biomarqueurs) et prédire la réponse probable d'un individu à un niveau d'exposition donné. Bien que ces deux approches soient extrêmement importantes, il convient de souligner qu'elles sont très différentes l'une de l'autre. Cet article portera sur la les facteurs génétiques susceptibilité individuelle sous-jacente à une exposition chimique particulière. Ce domaine de recherche est généralement appelé écogénétiqueou pharmacogénétique (voir Kalow 1962 et 1992). Bon nombre des progrès récents dans la détermination de la susceptibilité individuelle à la toxicité chimique ont évolué à partir d'une meilleure appréciation des processus par lesquels les humains et les autres mammifères détoxifient les produits chimiques et de la complexité remarquable des systèmes enzymatiques impliqués.
Figure 1. Interrelations entre l'évaluation de l'exposition, les différences ethniques, l'âge, le régime alimentaire, la nutrition et l'évaluation de la susceptibilité génétique - qui jouent tous un rôle dans le risque individuel de toxicité et de cancer
Nous décrirons d'abord la variabilité des réponses toxiques chez l'homme. Nous présenterons ensuite certaines des enzymes responsables d'une telle variation de réponse, due à des différences dans le métabolisme des produits chimiques étrangers. Ensuite, l'histoire et la nomenclature de la superfamille du cytochrome P450 seront détaillées. Cinq polymorphismes P450 humains ainsi que plusieurs polymorphismes non-P450 seront brièvement décrits ; ceux-ci sont responsables des différences humaines dans la réponse toxique. Nous discuterons ensuite d'un exemple pour souligner le fait que les différences génétiques chez les individus peuvent influencer l'évaluation de l'exposition, telle que déterminée par la surveillance environnementale. Enfin, nous discuterons du rôle de ces enzymes métabolisant les xénobiotiques dans les fonctions vitales critiques.
Variation de la réponse toxique parmi la population humaine
Les toxicologues et les pharmacologues parlent couramment de la dose létale moyenne pour 50 % de la population (LD50), la dose maximale moyenne tolérée pour 50 % de la population (MTD50), et la dose efficace moyenne d'un médicament particulier pour 50 % de la population (ED50). Cependant, comment ces doses affectent-elles chacun de nous sur une base individuelle ? En d'autres termes, un individu hautement sensible peut être 500 fois plus affecté ou 500 fois plus susceptible d'être affecté que l'individu le plus résistant d'une population ; pour ces personnes, le LD50 (et MDT50 et DE50) les valeurs auraient peu de sens. LD50, MDT50 et DE50 les valeurs ne sont pertinentes que lorsqu'elles se réfèrent à la population dans son ensemble.
Figure 2 illustre une relation dose-réponse hypothétique pour une réponse toxique par des individus dans une population donnée. Ce diagramme générique pourrait représenter le carcinome bronchique en réponse au nombre de cigarettes fumées, la chloracné en fonction des niveaux de dioxine sur le lieu de travail, l'asthme en fonction des concentrations d'ozone ou d'aldéhyde dans l'air, les coups de soleil en réponse à la lumière ultraviolette, la diminution du temps de coagulation en tant que une fonction de la prise d'aspirine, ou une détresse gastro-intestinale en réponse au nombre de jalapeno piments consommés. Généralement, dans chacun de ces cas, plus l'exposition est importante, plus la réponse toxique est importante. La plupart de la population présentera la moyenne et l'écart type de la réponse toxique en fonction de la dose. La « valeur aberrante résistante » (en bas à droite sur la figure 2) est un individu ayant moins de réponse à des doses ou des expositions plus élevées. Une « valeur aberrante sensible » (en haut à gauche) est un individu ayant une réponse exagérée à une dose ou une exposition relativement faible. Ces valeurs aberrantes, avec des différences extrêmes de réponse par rapport à la majorité des individus de la population, peuvent représenter des variantes génétiques importantes qui peuvent aider les scientifiques à tenter de comprendre les mécanismes moléculaires sous-jacents d'une réponse toxique.
Figure 2. Relation générique entre toute réponse toxique et la dose de tout agent environnemental, chimique ou physique
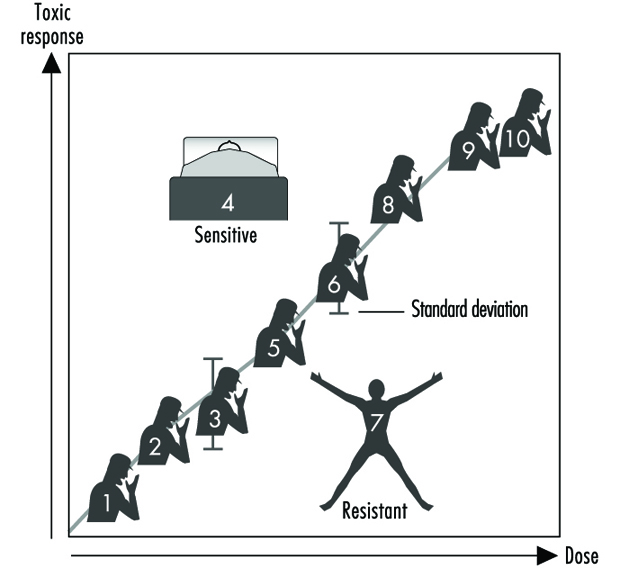
En utilisant ces valeurs aberrantes dans les études familiales, les scientifiques d'un certain nombre de laboratoires ont commencé à apprécier l'importance de l'hérédité mendélienne pour une réponse toxique donnée. Par la suite, on peut alors se tourner vers la biologie moléculaire et les études génétiques pour identifier le mécanisme sous-jacent au niveau du gène (génotype) responsable de la maladie causée par l'environnement (phénotype).
Enzymes xénobiotiques ou métabolisant les médicaments
Comment le corps réagit-il à la myriade de produits chimiques exogènes auxquels nous sommes exposés ? Les humains et d'autres mammifères ont développé des systèmes enzymatiques métaboliques très complexes comprenant plus d'une douzaine de superfamilles distinctes d'enzymes. Presque tous les produits chimiques auxquels les humains sont exposés seront modifiés par ces enzymes, afin de faciliter l'élimination de la substance étrangère du corps. Collectivement, ces enzymes sont fréquemment appelées enzymes métabolisant les médicaments or enzymes métabolisant les xénobiotiques. En fait, les deux termes sont des abus de langage. Premièrement, bon nombre de ces enzymes métabolisent non seulement des médicaments, mais aussi des centaines de milliers de produits chimiques environnementaux et alimentaires. Deuxièmement, toutes ces enzymes ont également des composés corporels normaux comme substrats ; aucune de ces enzymes ne métabolise uniquement les produits chimiques étrangers.
Depuis plus de quatre décennies, les processus métaboliques médiés par ces enzymes ont généralement été classés en réactions de phase I ou de phase II (figure 3). Les réactions de phase I («fonctionnalisation») impliquent généralement des modifications structurelles relativement mineures de la substance chimique mère par oxydation, réduction ou hydrolyse afin de produire un métabolite plus soluble dans l'eau. Fréquemment, les réactions de phase I fournissent une "manipulation" pour une modification supplémentaire d'un composé par des réactions de phase II ultérieures. Les réactions de phase I sont principalement médiées par une superfamille d'enzymes très polyvalentes, collectivement appelées cytochromes P450, bien que d'autres superfamilles d'enzymes puissent également être impliquées (figure 4).
Figure 3. Désignation classique des enzymes xénobiotiques ou métabolisant les médicaments de phase I et de phase II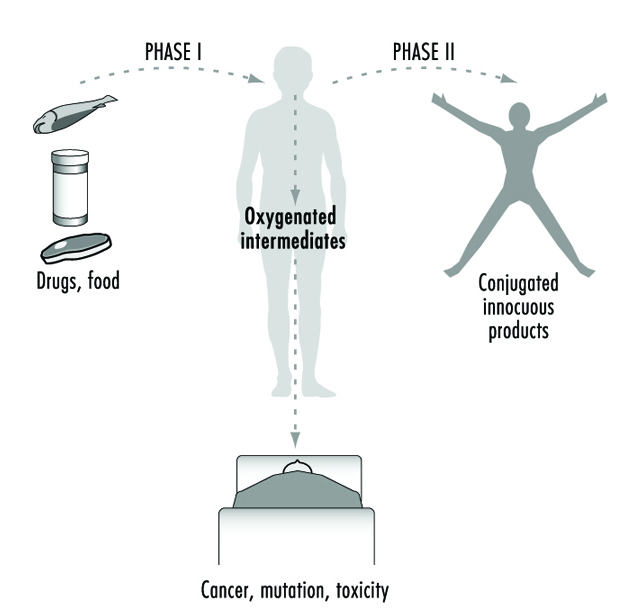
Figure 4. Exemples d'enzymes métabolisant les médicaments

Les réactions de phase II impliquent le couplage d'une molécule endogène soluble dans l'eau à un produit chimique (produit chimique parent ou métabolite de phase I) afin de faciliter l'excrétion. Les réactions de phase II sont fréquemment appelées réactions de « conjugaison » ou de « dérivatisation ». Les superfamilles d'enzymes catalysant les réactions de phase II sont généralement nommées en fonction de la fraction de conjugaison endogène impliquée : par exemple, l'acétylation par les N-acétyltransférases, la sulfatation par les sulfotransférases, la conjugaison du glutathion par les glutathion transférases et la glucuronidation par les UDP glucuronosyltransférases (figure 4). . Bien que le foie soit le principal organe du métabolisme des médicaments, les niveaux de certaines enzymes métabolisant les médicaments sont assez élevés dans le tractus gastro-intestinal, les gonades, les poumons, le cerveau et les reins, et ces enzymes sont sans aucun doute présentes dans une certaine mesure dans chaque cellule vivante.
Les enzymes métabolisant les xénobiotiques représentent un double tranchant Swords
Au fur et à mesure que nous en apprenons davantage sur les processus biologiques et chimiques conduisant à des aberrations pour la santé humaine, il est devenu de plus en plus évident que les enzymes métabolisant les médicaments fonctionnent de manière ambivalente (figure 3). Dans la majorité des cas, les produits chimiques liposolubles sont convertis en métabolites hydrosolubles plus facilement excrétés. Cependant, il est clair qu'en de nombreuses occasions, les mêmes enzymes sont capables de transformer d'autres produits chimiques inertes en molécules hautement réactives. Ces intermédiaires peuvent alors interagir avec des macromolécules cellulaires telles que les protéines et l'ADN. Ainsi, pour chaque produit chimique auquel les humains sont exposés, il existe un potentiel pour les voies concurrentes de activation métabolique ainsi que désintoxication.
Bref examen de la génétique
En génétique humaine, chaque gène (lieu) est situé sur l'une des 23 paires de chromosomes. Les deux allèles (un présent sur chaque chromosome de la paire) peuvent être identiques ou différents les uns des autres. Par exemple, le B ainsi que b allèles, dans lesquels B (yeux marrons) est dominant sur b (yeux bleus) : les individus de phénotype yeux bruns peuvent avoir soit BB or Bb génotypes, alors que les individus de phénotype aux yeux bleus ne peuvent avoir que bb génotype.
A polymorphisme est défini comme deux ou plusieurs phénotypes (traits) hérités de manière stable - dérivés du ou des mêmes gènes - qui sont maintenus dans la population, souvent pour des raisons pas nécessairement évidentes. Pour qu'un gène soit polymorphe, le produit du gène ne doit pas être essentiel au développement, à la vigueur reproductive ou à d'autres processus vitaux critiques. En fait, un « polymorphisme équilibré », dans lequel l'hétérozygote a un avantage de survie distinct sur l'un ou l'autre des homozygotes (par exemple, la résistance au paludisme et l'allèle drépanocytaire de l'hémoglobine) est une explication courante du maintien d'un allèle dans la population à un niveau élevé autrement inexpliqué. fréquences (voir Gonzalez et Nebert 1990).
Polymorphismes humains des enzymes métabolisant les xénobiotiques
Les différences génétiques dans le métabolisme de divers médicaments et produits chimiques environnementaux sont connues depuis plus de quatre décennies (Kalow 1962 et 1992). Ces différences sont fréquemment appelées pharmacogénétique ou, plus largement, polymorphismes écogénétiques. Ces polymorphismes représentent des allèles variants qui se produisent à une fréquence relativement élevée dans la population et sont généralement associés à des aberrations dans l'expression ou la fonction enzymatique. Historiquement, les polymorphismes étaient généralement identifiés suite à des réponses inattendues à des agents thérapeutiques. Plus récemment, la technologie de l'ADN recombinant a permis aux scientifiques d'identifier les altérations précises des gènes responsables de certains de ces polymorphismes. Les polymorphismes ont maintenant été caractérisés dans de nombreuses enzymes métabolisant les médicaments, y compris les enzymes de phase I et de phase II. Au fur et à mesure que de plus en plus de polymorphismes sont identifiés, il devient de plus en plus évident que chaque individu peut posséder un complément distinct d'enzymes métabolisant les médicaments. Cette diversité pourrait être qualifiée d'« empreinte métabolique ». C'est l'interaction complexe des diverses superfamilles d'enzymes métabolisant les médicaments au sein d'un individu qui déterminera finalement sa réponse particulière à un produit chimique donné (Kalow 1962 et 1992 ; Nebert 1988 ; Gonzalez et Nebert 1990 ; Nebert et Weber 1990).
Expression d'enzymes métabolisant les xénobiotiques humains dans la cellule Culture
Comment pourrions-nous développer de meilleurs prédicteurs des réponses toxiques humaines aux produits chimiques ? Les progrès dans la définition de la multiplicité des enzymes métabolisant les médicaments doivent s'accompagner d'une connaissance précise des enzymes qui déterminent le devenir métabolique des produits chimiques individuels. Les données recueillies dans les études de laboratoire sur les rongeurs ont certainement fourni des informations utiles. Cependant, des différences interspécifiques significatives dans les enzymes métabolisant les xénobiotiques nécessitent la prudence dans l'extrapolation des données aux populations humaines. Pour surmonter cette difficulté, de nombreux laboratoires ont développé des systèmes dans lesquels diverses lignées cellulaires en culture peuvent être modifiées pour produire des enzymes humaines fonctionnelles stables et à des concentrations élevées (Gonzalez, Crespi et Gelboin 1991). La production réussie d'enzymes humaines a été obtenue dans une variété de lignées cellulaires diverses provenant de sources comprenant des bactéries, des levures, des insectes et des mammifères.
Afin de définir encore plus précisément le métabolisme des substances chimiques, plusieurs enzymes ont également été produites avec succès dans une seule lignée cellulaire (Gonzalez, Crespi et Gelboin 1991). De telles lignées cellulaires fournissent des informations précieuses sur les enzymes précises impliquées dans le traitement métabolique d'un composé donné et de métabolites potentiellement toxiques. Si ces informations peuvent ensuite être combinées avec des connaissances concernant la présence et le niveau d'une enzyme dans les tissus humains, ces données devraient fournir des prédicteurs précieux de la réponse.
Cytochrome P450
Histoire et nomenclature
La superfamille du cytochrome P450 est l'une des superfamilles d'enzymes métabolisant les médicaments les plus étudiées, présentant une grande variabilité individuelle en réponse aux produits chimiques. Le cytochrome P450 est un terme générique pratique utilisé pour décrire une grande superfamille d'enzymes essentielles dans le métabolisme d'innombrables substrats endogènes et exogènes. Le terme cytochrome P450 a été inventé en 1962 pour décrire un inconnu pigment dans des cellules qui, lorsqu'elles sont réduites et liées avec du monoxyde de carbone, produisent un pic d'absorption caractéristique à 450 nm. Depuis le début des années 1980, la technologie de clonage d'ADNc a permis d'obtenir des informations remarquables sur la multiplicité des enzymes du cytochrome P450. À ce jour, plus de 400 gènes distincts du cytochrome P450 ont été identifiés chez les animaux, les plantes, les bactéries et les levures. Il a été estimé que n'importe quelle espèce de mammifère, comme les humains, peut posséder 60 gènes P450 distincts ou plus (Nebert et Nelson 1991). La multiplicité des gènes P450 a nécessité le développement d'un système de nomenclature standardisé (Nebert et al. 1987 ; Nelson et al. 1993). Proposé pour la première fois en 1987 et mis à jour sur une base semestrielle, le système de nomenclature est basé sur l'évolution divergente des comparaisons de séquences d'acides aminés entre les protéines P450. Les gènes P450 sont divisés en familles et sous-familles : les enzymes d'une famille présentent une similitude d'acides aminés supérieure à 40 %, et celles de la même sous-famille présentent une similitude de 55 %. Les gènes P450 sont nommés avec le symbole racine CYP suivi d'un chiffre arabe désignant la famille P450, d'une lettre désignant la sous-famille et d'un autre chiffre arabe désignant le gène individuel (Nelson et al. 1993; Nebert et al. 1991). Ainsi, CYP1A1 représente le gène P450 1 dans la famille 1 et la sous-famille A.
En février 1995, il y avait 403 CYP gènes dans la base de données, composée de 59 familles et 105 sous-familles. Celles-ci comprennent huit familles d'eucaryotes inférieurs, 15 familles de plantes et 19 familles de bactéries. Les 15 familles de gènes P450 humains comprennent 26 sous-familles, dont 22 ont été cartographiées à des emplacements chromosomiques dans la majeure partie du génome. Certaines séquences sont clairement orthologues dans de nombreuses espèces, par exemple, une seule CYP17 (stéroïde 17α-hydroxylase) a été trouvé chez tous les vertébrés examinés à ce jour ; d'autres séquences au sein d'une sous-famille sont fortement dupliquées, ce qui rend impossible l'identification de paires orthologues (par exemple, le CYP2C sous-famille). Fait intéressant, l'homme et la levure partagent un gène orthologue dans le CYP51 famille. De nombreuses revues complètes sont disponibles pour les lecteurs à la recherche d'informations complémentaires sur la superfamille P450 (Nelson et al. 1993 ; Nebert et al. 1991 ; Nebert et McKinnon 1994 ; Guengerich 1993 ; Gonzalez 1992).
Le succès du système de nomenclature P450 a entraîné le développement de systèmes terminologiques similaires pour les glucuronosyltransférases UDP (Burchell et al. 1991) et les mono-oxygénases contenant de la flavine (Lawton et al. 1994). Des systèmes de nomenclature similaires basés sur une évolution divergente sont également en cours de développement pour plusieurs autres superfamilles d'enzymes métabolisant les médicaments (par exemple, les sulfotransférases, les époxydes hydrolases et les aldéhydes déshydrogénases).
Récemment, nous avons divisé la superfamille des gènes P450 des mammifères en trois groupes (Nebert et McKinnon 1994) - ceux impliqués principalement dans le métabolisme chimique étranger, ceux impliqués dans la synthèse de diverses hormones stéroïdes et ceux participant à d'autres fonctions endogènes importantes. Ce sont les enzymes P450 métabolisant les xénobiotiques qui revêtent le plus d'importance pour la prédiction de la toxicité.
Enzymes P450 métabolisant les xénobiotiques
Les enzymes P450 impliquées dans le métabolisme des composés étrangers et des médicaments se trouvent presque toujours au sein des familles CYP1, CYP2, CYP3 ainsi que CYP4. Ces enzymes P450 catalysent une grande variété de réactions métaboliques, avec un seul P450 souvent capable de métaboliser de nombreux composés différents. De plus, plusieurs enzymes P450 peuvent métaboliser un seul composé à différents sites. En outre, un composé peut être métabolisé au même site unique par plusieurs P450, bien qu'à des vitesses variables.
Une propriété très importante des enzymes P450 métabolisant les médicaments est que nombre de ces gènes sont inductibles par les substances mêmes qui leur servent de substrats. D'autre part, d'autres gènes P450 sont induits par des non-substrats. Ce phénomène d'induction enzymatique est à la base de nombreuses interactions médicamenteuses d'importance thérapeutique.
Bien que présentes dans de nombreux tissus, ces enzymes P450 particulières se trouvent à des niveaux relativement élevés dans le foie, site principal du métabolisme des médicaments. Certaines des enzymes P450 métabolisant les xénobiotiques présentent une activité vis-à-vis de certains substrats endogènes (par exemple, l'acide arachidonique). Cependant, on pense généralement que la plupart de ces enzymes P450 métabolisant les xénobiotiques ne jouent pas de rôle physiologique important, bien que cela n'ait pas encore été établi expérimentalement. La perturbation homozygote sélective, ou "knock-out", des gènes P450 métabolisant les xénobiotiques individuels au moyen de méthodologies de ciblage génique chez la souris est susceptible de fournir bientôt des informations sans équivoque en ce qui concerne les rôles physiologiques des P450 métabolisant les xénobiotiques (pour une revue de ciblage génétique, voir Capecchi 1994).
Contrairement aux familles P450 codant pour les enzymes impliquées principalement dans les processus physiologiques, les familles codant pour les enzymes P450 métabolisant les xénobiotiques présentent une spécificité d'espèce marquée et contiennent fréquemment de nombreux gènes actifs par sous-famille (Nelson et al. 1993 ; Nebert et al. 1991). Compte tenu du manque apparent de substrats physiologiques, il est possible que les enzymes P450 des familles CYP1, CYP2, CYP3 ainsi que CYP4 qui sont apparus au cours des dernières centaines de millions d'années ont évolué comme un moyen de détoxification des produits chimiques étrangers rencontrés dans l'environnement et l'alimentation. De toute évidence, l'évolution des P450 métabolisant les xénobiotiques se serait produite sur une période qui précède de loin la synthèse de la plupart des produits chimiques synthétiques auxquels les humains sont maintenant exposés. Les gènes de ces quatre familles de gènes peuvent avoir évolué et divergé chez les animaux en raison de leur exposition aux métabolites des plantes au cours des 1.2 dernier milliard d'années - un processus appelé de manière descriptive « guerre animal-plante » (Gonzalez et Nebert 1990). La guerre animal-plante est le phénomène dans lequel les plantes ont développé de nouveaux produits chimiques (phytoalexines) comme mécanisme de défense afin d'empêcher l'ingestion par les animaux, et les animaux, à leur tour, ont répondu en développant de nouveaux gènes P450 pour s'adapter aux substrats diversifiés. Les exemples récemment décrits de guerre chimique plantes-insectes et plantes-champignons impliquant la détoxification P450 de substrats toxiques donnent un nouvel élan à cette proposition (Nebert 1994).
Ce qui suit est une brève introduction à plusieurs des polymorphismes de l'enzyme humaine P450 métabolisant les xénobiotiques dans lesquels on pense que les déterminants génétiques de la réponse toxique sont d'une grande importance. Jusqu'à récemment, les polymorphismes P450 étaient généralement suggérés par une variation inattendue de la réponse du patient aux agents thérapeutiques administrés. Plusieurs polymorphismes P450 sont en effet nommés en fonction du médicament avec lequel le polymorphisme a été identifié pour la première fois. Plus récemment, les efforts de recherche se sont concentrés sur l'identification des enzymes P450 précises impliquées dans le métabolisme des produits chimiques pour lesquels une variance est observée et la caractérisation précise des gènes P450 impliqués. Comme décrit précédemment, l'activité mesurable d'une enzyme P450 vis-à-vis d'un produit chimique modèle peut être appelée le phénotype. Les différences alléliques dans un gène P450 pour chaque individu sont appelées le génotype P450. Au fur et à mesure que l'analyse des gènes P450 est de plus en plus minutieuse, la base moléculaire précise de la variance phénotypique précédemment documentée devient plus claire.
La sous-famille CYP1A
Les CYP1A La sous-famille comprend deux enzymes chez l'homme et tous les autres mammifères : celles-ci sont désignées CYP1A1 et CYP1A2 sous la nomenclature standard P450. Ces enzymes présentent un intérêt considérable, car elles sont impliquées dans l'activation métabolique de nombreux procarcinogènes et sont également induites par plusieurs composés d'intérêt toxicologique, dont la dioxine. Par exemple, le CYP1A1 active métaboliquement de nombreux composés présents dans la fumée de cigarette. Le CYP1A2 active métaboliquement de nombreuses arylamines, associées au cancer de la vessie, trouvées dans l'industrie des colorants chimiques. Le CYP1A2 active également métaboliquement la 4-(méthylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK), une nitrosamine dérivée du tabac. CYP1A1 et CYP1A2 se trouvent également à des niveaux plus élevés dans les poumons des fumeurs de cigarettes, en raison de l'induction par les hydrocarbures polycycliques présents dans la fumée. Les niveaux d'activité du CYP1A1 et du CYP1A2 sont donc considérés comme des déterminants importants de la réponse individuelle à de nombreux produits chimiques potentiellement toxiques.
Intérêt toxicologique de la CYP1A sous-famille a été considérablement intensifiée par un rapport de 1973 établissant une corrélation entre le niveau d'inductibilité du CYP1A1 chez les fumeurs de cigarettes et la susceptibilité individuelle au cancer du poumon (Kellermann, Shaw et Luyten-Kellermann 1973). La base moléculaire de l'induction du CYP1A1 et du CYP1A2 a été au centre des préoccupations de nombreux laboratoires. Le processus d'induction est médié par une protéine appelée récepteur Ah à laquelle se lient les dioxines et les produits chimiques structurellement apparentés. Le nom Ah est dérivé de la aryl hnature hydrocarbonée de nombreux inducteurs du CYP1A. Fait intéressant, les différences dans le gène codant pour le récepteur Ah entre les souches de souris entraînent des différences marquées dans la réponse chimique et la toxicité. Un polymorphisme du gène du récepteur Ah semble également se produire chez l'homme : environ un dixième de la population présente une induction élevée du CYP1A1 et peut être plus à risque que les autres neuf dixièmes de la population de développer certains cancers induits chimiquement. Le rôle du récepteur Ah dans le contrôle des enzymes dans le CYP1A sous-famille, et son rôle en tant que déterminant de la réponse humaine à l'exposition chimique, a fait l'objet de plusieurs études récentes (Nebert, Petersen et Puga 1991; Nebert, Puga et Vasiliou 1993).
Existe-t-il d'autres polymorphismes qui pourraient contrôler le niveau de protéines CYP1A dans une cellule ? Un polymorphisme dans CYP1A1 a également été identifié, ce qui semble influencer le risque de cancer du poumon chez les fumeurs de cigarettes japonais, bien que ce même polymorphisme ne semble pas influencer le risque dans d'autres groupes ethniques (Nebert et McKinnon 1994).
CYP2C19
Les variations de la vitesse à laquelle les individus métabolisent le médicament anticonvulsivant (S)-méphénytoïne sont bien documentées depuis de nombreuses années (Guengerich 1989). Entre 2% et 5% des Caucasiens et jusqu'à 25% des Asiatiques sont déficients dans cette activité et peuvent être plus à risque de toxicité du médicament. On sait depuis longtemps que ce défaut enzymatique implique un membre de l'humain CYP2C sous-famille, mais la base moléculaire précise de cette déficience a fait l'objet d'une controverse considérable. La principale raison de cette difficulté était les six gènes ou plus dans le corps humain. CYP2C sous-famille. Cependant, il a été récemment démontré qu'une mutation à base unique dans le CYP2C19 est la principale cause de cette déficience (Goldstein et de Morais 1994). Un test ADN simple, basé sur la réaction en chaîne par polymérase (PCR), a également été développé pour identifier rapidement cette mutation dans les populations humaines (Goldstein et de Morais 1994).
CYP2D6
La variation la plus largement caractérisée dans un gène P450 est peut-être celle impliquant le CYP2D6 gène. Plus d'une dizaine d'exemples de mutations, réarrangements et délétions affectant ce gène ont été décrits (Meyer 1994). Ce polymorphisme a été suggéré pour la première fois il y a 20 ans par la variabilité clinique de la réponse des patients à l'agent antihypertenseur débrisoquine. Modifications dans le CYP2D6 gène donnant lieu à une activité enzymatique altérée sont donc collectivement appelés le polymorphisme débrisoquine.
Avant l'avènement des études basées sur l'ADN, les individus étaient classés comme métaboliseurs lents ou intensifs (PM, EM) de la débrisoquine sur la base des concentrations de métabolites dans les échantillons d'urine. Il est maintenant clair que les modifications de la CYP2D6 peut entraîner chez les individus non seulement un métabolisme médiocre ou étendu de la débrisoquine, mais également un métabolisme ultrarapide. La plupart des modifications dans le CYP2D6 gène sont associés à une déficience partielle ou totale de la fonction enzymatique ; cependant, des individus de deux familles ont récemment été décrits qui possèdent de multiples copies fonctionnelles du CYP2D6 gène, donnant lieu à un métabolisme ultrarapide des substrats du CYP2D6 (Meyer 1994). Cette observation remarquable fournit de nouvelles informations sur le large spectre d'activité du CYP2D6 précédemment observé dans les études de population. Les altérations de la fonction CYP2D6 revêtent une importance particulière, compte tenu des plus de 30 médicaments couramment prescrits métabolisés par cette enzyme. La fonction CYP2D6 d'un individu est donc un déterminant majeur de la réponse thérapeutique et toxique à la thérapie administrée. En effet, il a récemment été soutenu que la prise en compte du statut CYP2D6 d'un patient est nécessaire pour l'utilisation sûre des médicaments psychiatriques et cardiovasculaires.
Le rôle de l' CYP2D6 le polymorphisme en tant que déterminant de la susceptibilité individuelle aux maladies humaines telles que le cancer du poumon et la maladie de Parkinson a également fait l'objet d'études approfondies (Nebert et McKinnon 1994 ; Meyer 1994). Bien que les conclusions soient difficiles à définir compte tenu de la diversité des protocoles d'étude utilisés, la majorité des études semblent indiquer une association entre les métaboliseurs extensifs de la débrisoquine (phénotype EM) et le cancer du poumon. Les raisons d'une telle association ne sont pas claires pour le moment. Cependant, il a été démontré que l'enzyme CYP2D6 métabolise la NNK, une nitrosamine dérivée du tabac.
À mesure que les tests basés sur l'ADN s'améliorent, permettant une évaluation encore plus précise du statut du CYP2D6, il est prévu que la relation précise entre le CYP2D6 et le risque de maladie soit clarifiée. Alors que le métaboliseur rapide peut être lié à la susceptibilité au cancer du poumon, le métaboliseur lent (phénotype PM) semble être associé à la maladie de Parkinson de cause inconnue. Alors que ces études sont également difficiles à comparer, il semble que les individus PM ayant une capacité réduite à métaboliser les substrats du CYP2D6 (par exemple, la débrisoquine) ont un risque de développer la maladie de Parkinson de 2 à 2.5 fois.
CYP2E1
Les CYP2E1 Le gène code une enzyme qui métabolise de nombreux produits chimiques, y compris des médicaments et de nombreux cancérogènes de faible poids moléculaire. Cette enzyme est également intéressante car elle est hautement inductible par l'alcool et peut jouer un rôle dans les lésions hépatiques induites par des produits chimiques tels que le chloroforme, le chlorure de vinyle et le tétrachlorure de carbone. L'enzyme se trouve principalement dans le foie et le niveau d'enzyme varie considérablement d'un individu à l'autre. Un examen minutieux de la CYP2E1 a abouti à l'identification de plusieurs polymorphismes (Nebert et McKinnon 1994). Une relation a été rapportée entre la présence de certaines variations structurelles dans le CYP2E1 réduction génétique et apparente du risque de cancer du poumon dans certaines études ; cependant, il existe des différences interethniques claires qui nécessitent une clarification de cette relation possible.
La sous-famille CYP3A
Chez l'homme, quatre enzymes ont été identifiées comme membres de la CYP3A sous-famille en raison de leur similitude dans la séquence d'acides aminés. Les enzymes CYP3A métabolisent de nombreux médicaments couramment prescrits tels que l'érythromycine et la cyclosporine. L'aflatoxine B, contaminant alimentaire cancérigène1 est également un substrat du CYP3A. Un membre de l'humain CYP3A sous-famille, désignée CYP3A4, est le principal P450 dans le foie humain et est également présent dans le tractus gastro-intestinal. Comme c'est le cas pour de nombreuses autres enzymes P450, le niveau de CYP3A4 est très variable d'un individu à l'autre. Une deuxième enzyme, désignée CYP3A5, se trouve dans seulement environ 25 % des foies ; la base génétique de cette découverte n'a pas été élucidée. L'importance de la variabilité du CYP3A4 ou du CYP3A5 en tant que facteur dans les déterminants génétiques de la réponse toxique n'a pas encore été établie (Nebert et McKinnon, 1994).
Polymorphismes non P450
De nombreux polymorphismes existent également au sein d'autres superfamilles d'enzymes métabolisant les xénobiotiques (par exemple, les glutathion transférases, les UDP glucuronosyltransférases, les para-oxonases, les déshydrogénases, les N-acétyltransférases et les mono-oxygénases contenant de la flavine). Étant donné que la toxicité ultime de tout intermédiaire généré par P450 dépend de l'efficacité des réactions de détoxification de phase II ultérieures, le rôle combiné des polymorphismes enzymatiques multiples est important pour déterminer la sensibilité aux maladies induites chimiquement. L'équilibre métabolique entre les réactions de phase I et de phase II (figure 3) est donc susceptible d'être un facteur majeur dans les maladies humaines d'origine chimique et les déterminants génétiques de la réponse toxique.
Le polymorphisme du gène GSTM1
Un exemple bien étudié d'un polymorphisme dans une enzyme de phase II est celui impliquant un membre de la superfamille des enzymes glutathion S-transférase, désignée GST mu ou GSTM1. Cette enzyme particulière présente un intérêt toxicologique considérable car elle semble être impliquée dans la détoxification ultérieure des métabolites toxiques produits à partir de produits chimiques dans la fumée de cigarette par l'enzyme CYP1A1. Le polymorphisme identifié dans ce gène de la glutathion transférase implique une absence totale d'enzyme fonctionnelle chez jusqu'à la moitié de tous les Caucasiens étudiés. Cette absence d'enzyme de phase II semble être associée à une susceptibilité accrue au cancer du poumon. En regroupant les individus sur la base des deux variantes CYP1A1 gènes et la délétion ou la présence d'un GSTM1 gène, il a été démontré que le risque de développer un cancer du poumon induit par le tabagisme varie considérablement (Kawajiri, Watanabe et Hayashi 1994). En particulier, les individus présentant une rare CYP1A1 altération génétique, associée à une absence de GSTM1 , présentaient un risque plus élevé (jusqu'à neuf fois) de développer un cancer du poumon lorsqu'ils étaient exposés à un niveau relativement faible de fumée de cigarette. Fait intéressant, il semble y avoir des différences interethniques dans la signification des gènes variants qui nécessitent une étude plus approfondie afin d'élucider le rôle précis de ces altérations dans la susceptibilité à la maladie (Kalow 1962 ; Nebert et McKinnon 1994 ; Kawajiri, Watanabe et Hayashi 1994).
Effet synergique de deux ou plusieurs polymorphismes sur la toxicité réponse
Une réponse toxique à un agent environnemental peut être fortement exagérée par la combinaison de deux défauts pharmacogénétiques chez le même individu, par exemple, les effets combinés du polymorphisme de la N-acétyltransférase (NAT2) et du polymorphisme de la glucose-6-phosphate déshydrogénase (G6PD) .
L'exposition professionnelle aux arylamines constitue un risque grave de cancer de la vessie. Depuis les élégantes études de Cartwright en 1954, il est devenu clair que le statut de N-acétylateur est un déterminant du cancer de la vessie induit par les colorants azoïques. Il existe une corrélation hautement significative entre le phénotype acétyleur lent et la survenue d'un cancer de la vessie, ainsi que le degré d'envahissement de ce cancer dans la paroi vésicale. Au contraire, il existe une association significative entre le phénotype à acétylation rapide et l'incidence du carcinome colorectal. La N-acétyltransférase (NAT1, NAT2) ont été clonés et séquencés, et les tests basés sur l'ADN sont maintenant capables de détecter plus d'une douzaine de variantes alléliques qui expliquent le phénotype à acétyleur lent. Le NAT2 Le gène est polymorphe et responsable de la majeure partie de la variabilité de la réponse toxique aux produits chimiques environnementaux (Weber 1987; Grant 1993).
La glucose-6-phosphate déshydrogénase (G6PD) est une enzyme essentielle à la génération et au maintien du NADPH. Une activité G6PD faible ou absente peut entraîner une hémolyse sévère induite par des médicaments ou des xénobiotiques, en raison de l'absence de taux normaux de glutathion réduit (GSH) dans les globules rouges. Le déficit en G6PD touche au moins 300 millions de personnes dans le monde. Plus de 10% des hommes afro-américains présentent le phénotype le moins sévère, tandis que certaines communautés sardes présentent le «type méditerranéen» plus sévère à des fréquences pouvant atteindre une personne sur trois. Le G6PD Le gène a été cloné et localisé sur le chromosome X, et de nombreuses mutations ponctuelles diverses expliquent le degré élevé d'hétérogénéité phénotypique observée chez les individus déficients en G6PD (Beutler 1992).
La thiozalsulphone, une arylamine sulfamide, s'est avérée provoquer une distribution bimodale de l'anémie hémolytique dans la population traitée. Lorsqu'ils sont traités avec certains médicaments, les individus présentant la combinaison d'un déficit en G6PD et du phénotype d'acétylateur lent sont plus affectés que ceux présentant le déficit en G6PD seul ou le phénotype d'acétylateur lent seul. Les acétyleurs lents déficients en G6PD sont au moins 40 fois plus sensibles que les acétyleurs rapides normaux à la G6PD à l'hémolyse induite par la thiozalsulfone.
Effet des polymorphismes génétiques sur l'évaluation de l'exposition
L'évaluation de l'exposition et la biosurveillance (figure 1) nécessitent également des informations sur la constitution génétique de chaque individu. Étant donné une exposition identique à un produit chimique dangereux, le niveau d'adduits à l'hémoglobine (ou d'autres biomarqueurs) peut varier de deux ou trois ordres de grandeur entre les individus, en fonction de l'empreinte métabolique de chacun.
La même pharmacogénétique combinée a été étudiée chez des ouvriers d'usines chimiques en Allemagne (tableau 1). Les adduits d'hémoglobine chez les travailleurs exposés à l'aniline et à l'acétanilide sont de loin les plus élevés chez les acétyleurs lents déficients en G6PD, par rapport aux autres phénotypes pharmacogénétiques combinés possibles. Cette étude a des implications importantes pour l'évaluation de l'exposition. Ces données démontrent que, bien que deux personnes puissent être exposées au même niveau ambiant de produit chimique dangereux sur le lieu de travail, la quantité d'exposition (via des biomarqueurs tels que les adduits d'hémoglobine) peut être estimée à deux ordres de grandeur ou plus de moins, en raison à la prédisposition génétique sous-jacente de l'individu. De même, le risque résultant d'un effet néfaste sur la santé peut varier de deux ordres de grandeur ou plus.
Tableau 1 : Adduits à l'hémoglobine chez les travailleurs exposés à l'aniline et à l'acétanilide
| Statut d'acétylateur | Déficit en G6PD | |||
| Fast | Lent | Non | Oui | Adduits de l'Hb |
| + | + | 2 | ||
| + | + | 30 | ||
| + | + | 20 | ||
| + | + | 100 | ||
Source : Adapté de Lewalter et Korallus 1985.
Différences génétiques dans la liaison ainsi que dans le métabolisme
Il convient de souligner que le même cas fait ici pour le méta-bolisme peut également être fait pour la liaison. Les différences héréditaires dans la liaison des agents environnementaux affecteront grandement la réponse toxique. Par exemple, les différences dans la souris cdm peut profondément affecter la sensibilité individuelle à la nécrose testiculaire induite par le cadmium (Taylor, Heiniger et Meier 1973). Les différences d'affinité de liaison du récepteur Ah affectent probablement la toxicité induite par la dioxine et le cancer (Nebert, Petersen et Puga 1991; Nebert, Puga et Vasiliou 1993).
La figure 5 résume le rôle du métabolisme et de la liaison dans la toxicité et le cancer. Les agents toxiques, tels qu'ils existent dans l'environnement ou après le métabolisme ou la liaison, déclenchent leurs effets soit par une voie génotoxique (dans laquelle des dommages à l'ADN se produisent) soit par une voie non génotoxique (dans laquelle les dommages à l'ADN et la mutagenèse ne doivent pas se produire). Fait intéressant, il est récemment devenu clair que les agents endommageant l'ADN "classiques" peuvent fonctionner via une voie de transduction de signal non génotoxique dépendante du glutathion réduit (GSH), qui est initiée sur ou près de la surface cellulaire en l'absence d'ADN et à l'extérieur du noyau cellulaire. (Devary et al. 1993). Les différences génétiques dans le métabolisme et la liaison restent cependant les principaux déterminants du contrôle des différentes réponses toxiques individuelles.
Figure 5. Moyens généraux par lesquels la toxicité se produit

Rôle de la fonction cellulaire de l'enzyme métabolisant les médicaments
La variation génétique de la fonction enzymatique métabolisant les médicaments est d'une importance majeure pour déterminer la réponse individuelle aux produits chimiques. Ces enzymes sont essentielles pour déterminer le devenir et l'évolution dans le temps d'un produit chimique étranger après exposition.
Comme l'illustre la figure 5, l'importance des enzymes métabolisant les médicaments dans la sensibilité individuelle à l'exposition chimique peut en fait présenter un problème beaucoup plus complexe que ce qui ressort de cette simple discussion du métabolisme des xénobiotiques. En d'autres termes, au cours des deux dernières décennies, les mécanismes génotoxiques (mesures des adduits à l'ADN et des adduits aux protéines) ont été fortement mis en avant. Cependant, que se passe-t-il si les mécanismes non génotoxiques sont au moins aussi importants que les mécanismes génotoxiques pour provoquer des réponses toxiques ?
Comme mentionné précédemment, les rôles physiologiques de nombreuses enzymes métabolisant les médicaments impliquées dans le métabolisme des xénobiotiques n'ont pas été définis avec précision. Nebert (1994) a proposé qu'en raison de leur présence sur cette planète depuis plus de 3.5 milliards d'années, les enzymes métabolisant les médicaments étaient à l'origine (et sont encore principalement) responsables de la régulation des niveaux cellulaires de nombreux ligands non peptidiques importants dans l'activation transcriptionnelle. de gènes affectant la croissance, la différenciation, l'apoptose, l'homéostasie et les fonctions neuroendocrines. De plus, la toxicité de la plupart, sinon de la totalité, des agents environnementaux se produit au moyen de Agoniste or antagoniste action sur ces voies de transduction du signal (Nebert 1994). Sur la base de cette hypothèse, la variabilité génétique des enzymes métabolisant les médicaments peut avoir des effets assez dramatiques sur de nombreux processus biochimiques critiques au sein de la cellule, entraînant ainsi des différences importantes dans la réponse toxique. Il est en effet possible qu'un tel scénario puisse également être à l'origine de nombreux effets indésirables idiosyncratiques rencontrés chez les patients utilisant des médicaments couramment prescrits.
Conclusions
La dernière décennie a vu des progrès remarquables dans notre compréhension de la base génétique de la réponse différentielle aux produits chimiques dans les médicaments, les aliments et les polluants environnementaux. Les enzymes métabolisant les médicaments ont une profonde influence sur la façon dont les humains réagissent aux produits chimiques. À mesure que notre connaissance de la multiplicité des enzymes métabolisant les médicaments continue d'évoluer, nous sommes de plus en plus en mesure d'améliorer les évaluations du risque toxique pour de nombreux médicaments et produits chimiques environnementaux. Ceci est peut-être le plus clairement illustré dans le cas de l'enzyme CYP2D6 du cytochrome P450. En utilisant des tests basés sur l'ADN relativement simples, il est possible de prédire la réponse probable de tout médicament principalement métabolisé par cette enzyme ; cette prédiction garantira une utilisation plus sûre de médicaments précieux, mais potentiellement toxiques.
L'avenir verra sans aucun doute une explosion dans l'identification d'autres polymorphismes (phénotypes) impliquant des enzymes métabolisant les médicaments. Ces informations seront accompagnées de tests ADN améliorés et peu invasifs pour identifier les génotypes dans les populations humaines.
De telles études devraient être particulièrement instructives pour évaluer le rôle des produits chimiques dans les nombreuses maladies environnementales d'origine actuellement inconnue. La prise en compte de multiples polymorphismes d'enzymes métabolisant les médicaments, en combinaison (par exemple, tableau 1), est également susceptible de représenter un domaine de recherche particulièrement fertile. De telles études permettront de clarifier le rôle des produits chimiques dans l'étiologie des cancers. Collectivement, ces informations devraient permettre de formuler des conseils de plus en plus individualisés sur l'évitement des produits chimiques susceptibles d'être une préoccupation individuelle. C'est le domaine de la toxicologie préventive. De tels conseils aideront sans aucun doute grandement tous les individus à faire face à la charge chimique toujours croissante à laquelle nous sommes exposés.