Генетска токсикологија, по дефиницији, је студија о томе како хемијски или физички агенси утичу на замршени процес наслеђа. Генотоксичне хемикалије се дефинишу као једињења која су способна да модификују наследни материјал живих ћелија. Вероватноћа да ће одређена хемикалија изазвати генетско оштећење неизбежно зависи од неколико варијабли, укључујући ниво изложености организма хемикалији, дистрибуцију и задржавање хемикалије када уђе у тело, ефикасност метаболичке активације и/или система детоксикације у циљна ткива, и реактивност хемикалије или њених метаболита са критичним макромолекулима унутар ћелија. Вероватноћа да ће генетско оштећење изазвати болест на крају зависи од природе оштећења, способности ћелије да поправи или појача генетско оштећење, могућности да изрази било коју промену која је изазвана, и способности тела да препозна и потисне умножавање аберантне ћелије.
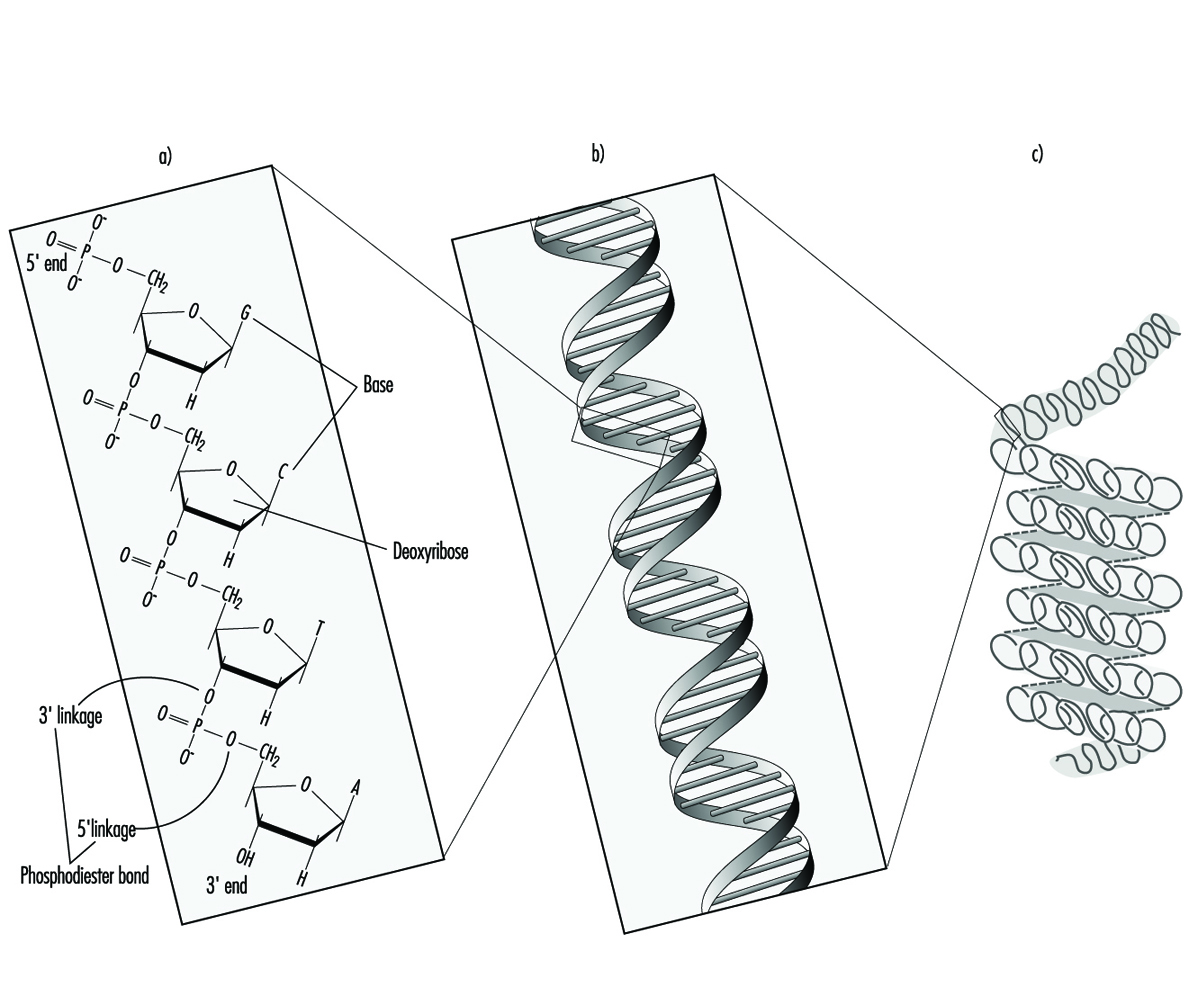
У вишим организмима, наследне информације су организоване у хромозомима. Хромозоми се састоје од чврсто кондензованих ланаца ДНК повезане са протеинима. У оквиру једног хромозома, сваки молекул ДНК постоји као пар дугих, неразгранатих ланаца нуклеотидних подјединица повезаних фосфодиестарским везама које спајају 5 угљеника једног дела дезоксирибозе са 3 угљеника следећег (слика 1). Поред тога, једна од четири различите нуклеотидне базе (аденин, цитозин, гванин или тимин) је везана за сваку подјединицу дезоксирибозе као перле на нити. Тродимензионално, сваки пар ланаца ДНК формира двоструку спиралу са свим базама оријентисаним ка унутрашњости спирале. Унутар хеликса, свака база је повезана са својом комплементарном базом на супротном ДНК ланцу; водонична веза диктира снажно, нековалентно упаривање аденина са тимином и гванина са цитозином (слика 1). Пошто је секвенца нуклеотидних база комплементарна целом дужином молекула дуплекс ДНК, оба ланца носе у суштини исте генетске информације. У ствари, током репликације ДНК сваки ланац служи као шаблон за производњу новог партнерског ланца.
Слика 1. (а) примарна, (б) секундарна и (ц) терцијарна организација људских наследних информација
 Користећи РНК и низ различитих протеина, ћелија на крају дешифрује информације кодиране линеарном секвенцом база унутар специфичних региона ДНК (гена) и производи протеине који су неопходни за опстанак основних ћелија, као и за нормалан раст и диференцијацију. У суштини, нуклеотиди функционишу као биолошка абецеда која се користи за кодирање аминокиселина, градивних блокова протеина.
Користећи РНК и низ различитих протеина, ћелија на крају дешифрује информације кодиране линеарном секвенцом база унутар специфичних региона ДНК (гена) и производи протеине који су неопходни за опстанак основних ћелија, као и за нормалан раст и диференцијацију. У суштини, нуклеотиди функционишу као биолошка абецеда која се користи за кодирање аминокиселина, градивних блокова протеина.
Када се уметну нетачни нуклеотиди или се нуклеотиди изгубе, или када се додају непотребни нуклеотиди током синтезе ДНК, грешка се назива мутација. Процењено је да се на сваких 10 јавља мање од једне мутације9 нуклеотиди уграђени током нормалне репликације ћелија. Иако мутације нису нужно штетне, промене које изазивају инактивацију или прекомерну експресију важних гена могу довести до разних поремећаја, укључујући рак, наследне болести, развојне абнормалности, неплодност и ембрионалну или перинаталну смрт. Веома ретко, мутација може довести до побољшаног преживљавања; такве појаве су основа природне селекције.
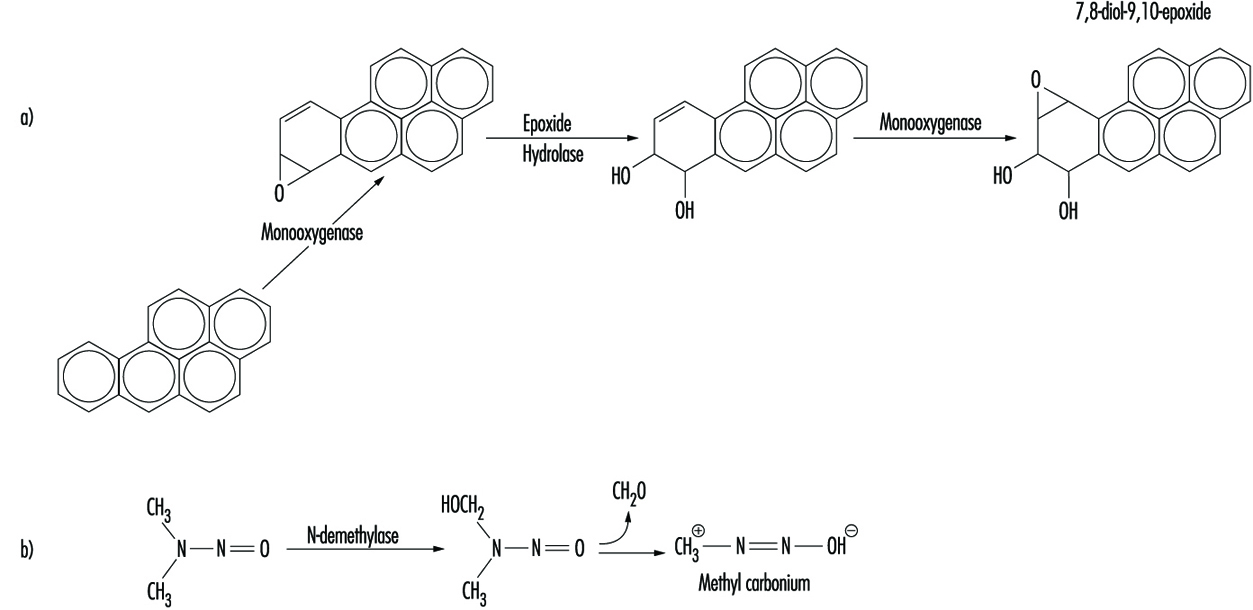
Иако неке хемикалије реагују директно са ДНК, већина захтева метаболичку активацију. У последњем случају, електрофилни интермедијери као што су епоксиди или јони угљеника су на крају одговорни за изазивање лезија на различитим нуклеофилним местима унутар генетског материјала (слика 2). У другим случајевима, генотоксичност је посредована нуспроизводима интеракције једињења са интрацелуларним липидима, протеинима или кисеоником.
Слика 2. Биоактивација: а) бензо(а)пирена; и б) Н-нитрозодиметиламин
Због њиховог релативног обиља у ћелијама, протеини су најчешћа мета интеракције токсичних супстанци. Међутим, модификација ДНК изазива већу забринутост због централне улоге овог молекула у регулисању раста и диференцијације кроз више генерација ћелија.
На молекуларном нивоу, електрофилна једињења имају тенденцију да нападају кисеоник и азот у ДНК. Локације које су најсклоне модификацијама илустроване су на слици 3. Иако су кисеоник унутар фосфатних група у ДНК кичми такође мета за хемијску модификацију, сматра се да је оштећење база биолошки релевантније јер се ове групе сматрају примарним информативним елемената у молекулу ДНК.
Слика 3. Примарна места хемијски изазваних оштећења ДНК

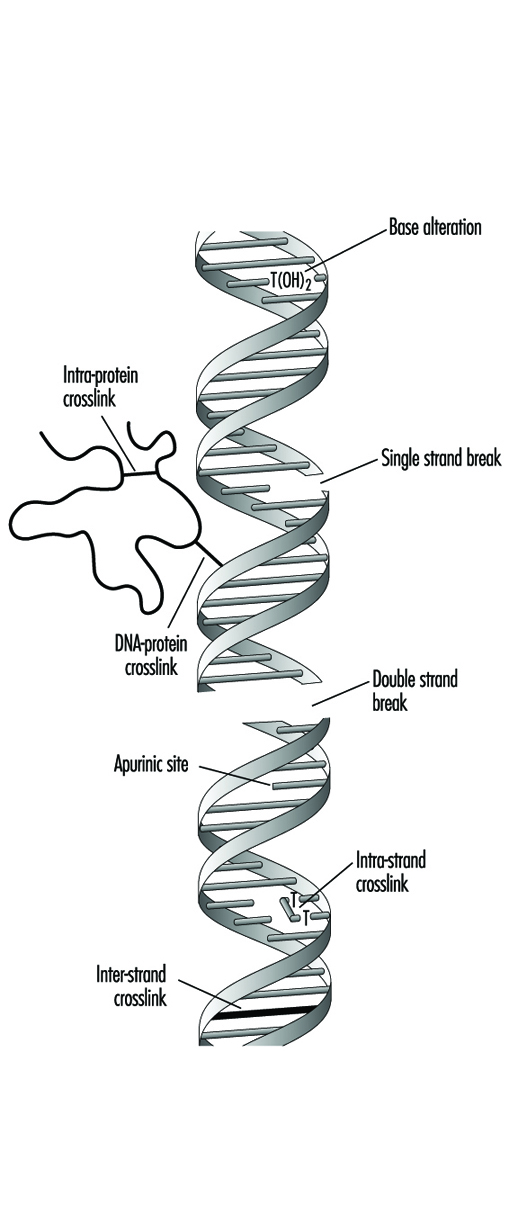
Једињења која садрже један електрофилни део обично испољавају генотоксичност тако што производе моно-адукте у ДНК. Слично томе, једињења која садрже два или више реактивних делова могу да реагују са два различита нуклеофилна центра и на тај начин произведу интра- или интер-молекуларне попречне везе у генетском материјалу (слика 4). Унакрсне везе између ДНК-ДНК и ДНК-протеина могу бити посебно цитотоксичне јер могу формирати потпуне блокове за репликацију ДНК. Из очигледних разлога, смрт ћелије елиминише могућност да ће бити мутирана или неопластично трансформисана. Генотоксични агенси такође могу деловати тако што изазивају прекиде у фосфодиестарској кичми, или између база и шећера (производећи абазична места) у ДНК. Такви прекиди могу бити директан резултат хемијске реактивности на месту оштећења или се могу јавити током поправке једног од горе наведених типова лезија ДНК.
Слика 4. Разне врсте оштећења комплекса протеин-ДНК
Током протеклих тридесет до четрдесет година развијене су различите технике за праћење врсте генетских оштећења изазваних разним хемикалијама. Такви тестови су детаљно описани на другом месту у овом поглављу и Енциклопедија.
Погрешна репликација „микролезија“ као што су моно-адукти, абазична места или прекиди једног ланца може на крају довести до супституција нуклеотидних базних парова, или уметања или брисања кратких полинуклеотидних фрагмената у хромозомској ДНК. Насупрот томе, „макролезије“, као што су гломазни адукти, унакрсне везе или прекиди двоструког ланца, могу изазвати добијање, губитак или преуређење релативно великих делова хромозома. У сваком случају, последице могу бити погубне по организам јер било који од ових догађаја може довести до смрти ћелије, губитка функције или малигне трансформације ћелија. Како тачно оштећење ДНК узрокује рак је углавном непознато. Тренутно се верује да процес може укључивати неодговарајућу активацију прото-онкогена као што је нпр миц Рас, и/или инактивација недавно идентификованих гена супресора тумора као што је п53. Абнормална експресија било ког типа гена укида нормалне ћелијске механизме за контролу пролиферације и/или диференцијације ћелија.
Превласт експерименталних доказа указује да је развој канцера након излагања електрофилним једињењима релативно редак догађај. Ово се делимично може објаснити интринзичном способношћу ћелије да препозна и поправи оштећену ДНК или неуспехом ћелија са оштећеном ДНК да преживе. Током поправке, оштећена база, нуклеотид или кратак део нуклеотида који окружује место оштећења се уклања и (користећи супротни ланац као шаблон) нови део ДНК се синтетише и спаја на место. Да би била ефикасна, поправка ДНК мора да се деси са великом тачношћу пре деобе ћелије, пре него што дође до могућности за ширење мутације.
Клиничке студије су показале да људи са наследним дефектима у способности да поправе оштећену ДНК често развију рак и/или развојне абнормалности у раном добу (табела 1). Такви примери пружају снажне доказе који повезују акумулацију оштећења ДНК са људским болестима. Слично, агенси који промовишу пролиферацију ћелија (као што је тетрадеканоилфорбол ацетат) често појачавају карциногенезу. За ова једињења, повећана вероватноћа неопластичне трансформације може бити директна последица смањења времена доступног ћелији да изврши адекватну поправку ДНК.
Табела 1. Наследни поремећаји склони карциному за које се чини да укључују дефекте у поправци ДНК
| Синдром | simptomi | Ћелијски фенотип |
| Атаксија телангиектазија | Неуролошко погоршање Иммунодефициенци Висока инциденција лимфома |
Преосетљивост на јонизујуће зрачење и одређене алкилирајуће агенсе. Дисрегулисана репликација оштећене ДНК (може указивати на скраћено време за поправку ДНК) |
| Блоомов синдром | Абнормалности у развоју Лезије на изложеној кожи Висока инциденција тумора имуног система и гастроинтестиналног тракта |
Висока учесталост хромозомских аберација Дефектна лигација прекида повезаних са поправком ДНК |
| Фанцонијева анемија | Заостајање у расту Висока учесталост леукемије |
Преосетљивост на средства за умрежавање Висока учесталост хромозомских аберација Дефектна поправка унакрсних веза у ДНК |
| Наследни неполипозни рак дебелог црева | Висока учесталост рака дебелог црева | Дефект у поправци неусклађености ДНК (када се убаци погрешан нуклеотид током репликације) |
| Ксеродерма пигментосум | Висока инциденца епителиома на изложеним деловима коже Неуролошко оштећење (у многим случајевима) |
Преосетљивост на УВ светлост и многе хемијске карциногене Дефекти у поправци ексцизије и/или репликацији оштећене ДНК |
Најраније теорије о томе како хемикалије интерагују са ДНК могу се пратити до студија спроведених током развоја иперита за употребу у рату. Даље разумевање је произашло из напора да се идентификују агенси против рака који би селективно зауставили репликацију туморских ћелија које се брзо деле. Повећана забринутост јавности због опасности у нашем окружењу подстакла је додатна истраживања механизама и последица хемијске интеракције са генетским материјалом. Примери различитих врста хемикалија које испољавају генотоксичност су представљени у табели 2.
Табела 2. Примери хемикалија које показују генотоксичност у људским ћелијама
| Класа хемикалија | Пример | Извор излагања | Вероватна генотоксична лезија |
| Афлатоксини | Афлатоксин Б1 | Контаминирана храна | Крупни ДНК адукти |
| Ароматични амини | 2-Ацетиламинофлуорен | еколошки | Крупни ДНК адукти |
| Азиридин кинони | Митомицин Ц | Хемотерапија рака | Моно-адукти, међуланчане унакрсне везе и једноланчани прекиди у ДНК. |
| Хлоровани угљоводоници | Винил хлорид | еколошки | Моно-адукти у ДНК |
| Метали и једињења метала | Цисплатин | Хемотерапија рака | Унакрсне везе унутар и међу ланцима у ДНК |
| Једињења никла | еколошки | Моно-адукти и једноланчани прекиди у ДНК | |
| Азотни сенф | Циклофосфамид | Хемотерапија рака | Моно-адукти и међуланчане умрежене везе у ДНК |
| Нитросамини | Н-нитрозодиметиламин | Контаминирана храна | Моно-адукти у ДНК |
| Полицикличних ароматичних угљоводоника | Бензо (а) пирене | еколошки | Крупни ДНК адукти |