Anatomy
The ear is the sensory organ responsible for hearing and the maintenance of equilibrium, via the detection of body position and of head movement. It is composed of three parts: the outer, middle, and inner ear; the outer ear lies outside the skull, while the other two parts are embedded in the temporal bone (figure 1).
Figure 1. Diagram of the ear.

The outer ear consists of the auricle, a cartilaginous skin-covered structure, and the external auditory canal, an irregularly-shaped cylinder approximately 25 mm long which is lined by glands secreting wax.
The middle ear consists of the tympanic cavity, an air-filled cavity whose outer walls form the tympanic membrane (eardrum), and communicates proximally with the nasopharynx by the Eustachian tubes, which maintain pressure equilibrium on either side of the tympanic membrane. For instance, this communication explains how swallowing allows equalization of pressure and restoration of lost hearing acuity caused by rapid change in barometric pressure (e.g., landing airplanes, fast elevators). The tympanic cavity also contains the ossicles—the malleus, incus and stapes—which are controlled by the stapedius and tensor tympani muscles. The tympanic membrane is linked to the inner ear by the ossicles, specifically by the mobile foot of the stapes, which lies against the oval window.
The inner ear contains the sensory apparatus per se. It consists of a bony shell (the bony labyrinth) within which is found the membranous labyrinth—a series of cavities forming a closed system filled with endolymph, a potassium-rich liquid. The membranous labyrinth is separated from the bony labyrinth by the perilymph, a sodium-rich liquid.
The bony labyrinth itself is composed of two parts. The anterior portion is known as the cochlea and is the actual organ of hearing. It has a spiral shape reminiscent of a snail shell, and is pointed in the anterior direction. The posterior portion of the bony labyrinth contains the vestibule and the semicircular canals, and is responsible for equilibrium. The neurosensory structures involved in hearing and equilibrium are located in the membranous labyrinth: the organ of Corti is located in the cochlear canal, while the maculae of the utricle and the saccule and the ampullae of the semicircular canals are located in the posterior section.
Hearing organs
The cochlear canal is a spiral triangular tube, comprising two and one-half turns, which separates the scala vestibuli from the scala tympani. One end terminates in the spiral ligament, a process of the cochlea’s central column, while the other is connected to the bony wall of the cochlea.
The scala vestibuli and tympani end in the oval window (the foot of the stapes) and round window, respectively. The two chambers communicate through the helicotrema, the tip of the cochlea. The basilar membrane forms the inferior surface of the cochlear canal, and supports the organ of Corti, responsible for the transduction of acoustic stimuli. All auditory information is transduced by only 15,000 hair cells (organ of Corti), of which the so-called inner hair cells, numbering 3,500, are critically important, since they form synapses with approximately 90% of the 30,000 primary auditory neurons (figure 2). The inner and outer hair cells are separated from each other by an abundant layer of support cells. Traversing an extraordinarily thin membrane, the cilia of the hair cells are embedded in the tectorial membrane, whose free end is located above the cells. The superior surface of the cochlear canal is formed by Reissner’s membrane.
Figure 2. Cross-section of one loop of the cochlea. Diameter: approximately 1.5 mm.
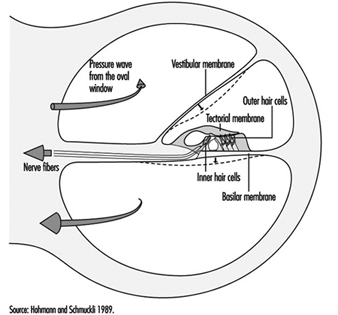
The bodies of the cochlear sensory cells resting on the basilar membrane are surrounded by nerve terminals, and their approximately 30,000 axons form the cochlear nerve. The cochlear nerve crosses the inner ear canal and extends to the central structures of the brain stem, the oldest part of the brain. The auditory fibres end their tortuous path in the temporal lobe, the part of the cerebral cortex responsible for the perception of acoustic stimuli.
Organs of Equilibrium
The sensory cells are located in the ampullae of the semicircular canals and the maculae of the utricle and saccule, and are stimulated by pressure transmitted through the endolymph as a result of head or body movements. The cells connect with bipolar cells whose peripheral processes form two tracts, one from the anterior and external semicircular canals, the other from the posterior semicircular canal. These two tracts enter the inner ear canal and unite to form the vestibular nerve, which extends to the vestibular nuclei in the brainstem. Fibres from the vestibular nuclei, in turn, extend to cerebellar centres controlling eye movements, and to the spinal cord.
The union of the vestibular and cochlear nerves forms the 8th cranial nerve, also known as the vestibulocochlear nerve.
Physiology of Hearing
Sound conduction through air
The ear is composed of a sound conductor (the outer and middle ear) and a sound receptor (the inner ear).
Sound waves passing through the external auditory canal strike the tympanic membrane, causing it to vibrate. This vibration is transmitted to the stapes through the hammer and anvil. The surface area of the tympanic membrane is almost 16 times that of the foot of the stapes (55 mm2/3.5 mm2), and this, in combination with the lever mechanism of the ossicles, results in a 22-fold amplification of the sound pressure. Due to the middle ear’s resonant frequency, the transmission ratio is optimal between 1,000 and 2,000 Hz. As the foot of the stapes moves, it causes waves to form in the liquid within the vestibular canal. Since the liquid is incompressible, each inward movement of the foot of the stapes causes an equivalent outward movement of the round window, towards the middle ear.
When exposed to high sound levels, the stapes muscle contracts, protecting the inner ear (the attenuation reflex). In addition to this function, the muscles of the middle ear also extend the dynamic range of the ear, improve sound localization, reduce resonance in the middle ear, and control air pressure in the middle ear and liquid pressure in the inner ear.
Between 250 and 4,000 Hz, the threshold of the attenuation reflex is approximately 80 decibels (dB) above the hearing threshold, and increases by approximately 0.6 dB/dB as the stimulation intensity increases. Its latency is 150 ms at threshold, and 24-35 ms in the presence of intense stimuli. At frequencies below the natural resonance of the middle ear, contraction of the middle ear muscles attenuates sound transmission by approximately 10 dB. Because of its latency, the attenuation reflex provides adequate protection from noise generated at rates above two to three per second, but not from discrete impulse noise.
The speed with which sound waves propagate through the ear depends on the elasticity of the basilar membrane. The elasticity increases, and the wave velocity thus decreases, from the base of the cochlea to the tip. The transfer of vibration energy to Reissner’s membrane and the basilar membrane is frequency-dependent. At high frequencies, the wave amplitude is greatest at the base, while for lower frequencies, it is greatest at the tip. Thus, the point of greatest mechanical excitation in the cochlea is frequency-dependent. This phenomenon underlies the ability to detect frequency differences. Movement of the basilar membrane induces shear forces in the stereocilia of the hair cells and triggers a series of mechanical, electrical and biochemical events responsible for mechanical-sensory transduction and initial acoustic signal processing. The shear forces on the stereocilia cause ionic channels in the cell membranes to open, modifying the permeability of the membranes and allowing the entry of potassium ions into the cells. This influx of potassium ions results in depolarization and the generation of an action potential.
Neurotransmitters liberated at the synaptic junction of the inner hair cells as a result of depolarization trigger neuronal impulses which travel down the afferent fibres of the auditory nerve toward higher centres. The intensity of auditory stimulation depends on the number of action potentials per unit time and the number of cells stimulated, while the perceived frequency of the sound depends on the specific nerve fibre populations activated. There is a specific spatial mapping between the frequency of the sound stimulus and the section of the cerebral cortex stimulated.
The inner hair cells are mechanoreceptors which transform signals generated in response to acoustic vibration into electric messages sent to the central nervous system. They are not, however, responsible for the ear’s threshold sensitivity and its extraordinary frequency selectivity.
The outer hair cells, on the other hand, send no auditory signals to the brain. Rather, their function is to selectively amplify mechano-acoustic vibration at near-threshold levels by a factor of approximately 100 (i.e., 40 dB), and so facilitate stimulation of inner hair cells. This amplification is believed to function through micromechanical coupling involving the tectorial membrane. The outer hair cells can produce more energy than they receive from external stimuli and, by contracting actively at very high frequencies, can function as cochlear amplifiers.
In the inner ear, interference between outer and inner hair cells creates a feedback loop which permits control of auditory reception, particularly of threshold sensitivity and frequency selectivity. Efferent cochlear fibres may thus help reduce cochlear damage caused by exposure to intense acoustic stimuli. Outer hair cells may also undergo reflex contraction in the presence of intense stimuli. The attenuation reflex of the middle ear, active primarily at low frequencies, and the reflex contraction in the inner ear, active at high frequencies, are thus complementary.
Bone conduction of sound
Sound waves may also be transmitted through the skull. Two mechanisms are possible:
In the first, compression waves impacting the skull cause the incompressible perilymph to deform the round or oval window. As the two windows have differing elasticities, movement of the endolymph results in movement of the basilar membrane.
The second mechanism is based on the fact that movement of the ossicles induces movement in the scala vestibuli only. In this mechanism, movement of the basilar membrane results from the translational movement produced by the inertia.
Bone conduction is normally 30-50 dB lower than air conduction—as is readily apparent when both ears are blocked. This is only true, however, for air-mediated stimuli, direct bone stimulation being attenuated to a different degree.
Sensitivity range
Mechanical vibration induces potential changes in the cells of the inner ear, conduction pathways and higher centres. Only frequencies of 16 Hz–25,000 Hz and sound pressures (these can be expressed in pascals, Pa) of 20 μPa to 20 Pa can be perceived. The range of sound pressures which can be perceived is remarkable—a 1-million-fold range! The detection thresholds of sound pressure are frequency-dependent, lowest at 1,000-6,000 Hz and increasing at both higher and lower frequencies.
For practical purposes, the sound pressure level is expressed in decibels (dB), a logarithmic measurement scale corresponding to perceived sound intensity relative to the auditory threshold. Thus, 20 μPa is equivalent to 0 dB. As the sound pressure increases tenfold, the decibel level increases by 20 dB, in accordance with the following formula:
Lx = 20 log Px/P0
where:
Lx = sound pressure in dB
Px = sound pressure in pascals
P0 = reference sound pressure(2×10–5 Pa, the auditory threshold)
The frequency-discrimination threshold, that is the minimal detectable difference in frequency, is 1.5 Hz up to 500 Hz, and 0.3% of the stimulus frequency at higher frequencies. At sound pressures near the auditory threshold, the sound-pressure-discrimination threshold is approximately 20%, although differences of as little as 2% may be detected at high sound pressures.
If two sounds differ in frequency by a sufficiently small amount, only one tone will be heard. The perceived frequency of the tone will be midway between the two source tones, but its sound pressure level is variable. If two acoustic stimuli have similar frequencies but differing intensities, a masking effect occurs. If the difference in sound pressure is large enough, masking will be complete, with only the loudest sound perceived.
Localization of acoustic stimuli depends on the detection of the time lag between the arrival of the stimulus at each ear, and, as such, requires intact bilateral hearing. The smallest detectable time lag is 3 x 10–5 seconds. Localization is facilitated by the head’s screening effect, which results in differences in stimulus intensity at each ear.
The remarkable ability of human beings to resolve acoustic stimuli is a result of frequency decomposition by the inner ear and frequency analysis by the brain. These are the mechanisms that allow individual sound sources such as individual musical instruments to be detected and identified in the complex acoustic signals that make up the music of a full symphony orchestra.
Physiopathology
Ciliary damage
The ciliary motion induced by intense acoustic stimuli may exceed the mechanical resistance of the cilia and cause mechanical destruction of hair cells. As these cells are limited in number and incapable of regeneration, any cell loss is permanent, and if exposure to the harmful sound stimulus continues, progressive. In general, the ultimate effect of ciliary damage is the development of a hearing deficit.
Outer hair cells are the most sensitive cells to sound and toxic agents such as anoxia, ototoxic medications and chemicals (e.g., quinine derivates, streptomycin and some other antibiotics, some anti-tumour preparations), and are thus the first to be lost. Only passive hydromechanical phenomena remain operative in outer hair cells which are damaged or have damaged stereocilia. Under these conditions, only gross analysis of acoustic vibration is possible. In very rough terms, cilia destruction in outer hair cells results in a 40 dB increase in hearing threshold.
Cellular damage
Exposure to noise, especially if it is repetitive or prolonged, may also affect the metabolism of cells of the organ of Corti, and afferent synapses located beneath the inner hair cells. Reported extraciliary effects include modification of cell ultrastructure (reticulum, mitochondria, lysosomes) and, postsynaptically, swelling of afferent dendrites. Dendritic swelling is probably due to the toxic accumulation of neurotransmitters as a result of excessive activity by inner hair cells. Nevertheless, the extent of stereociliary damage appears to determine whether hearing loss is temporary or permanent.
Noise-induced Hearing Loss
Noise is a serious hazard to hearing in today’s increasingly complex industrial societies. For example, noise exposure accounts for approximately one-third of the 28 million cases of hearing loss in the United States, and NIOSH (the National Institute for Occupational Safety and Health) reports that 14% of American workers are exposed to potentially dangerous sound levels, that is levels exceeding 90 dB. Noise exposure is the most widespread harmful occupational exposure and is the second leading cause, after age-related effects, of hearing loss. Finally, the contribution of non-occupational noise exposure must not be forgotten, such as home workshops, over-amplified music especially with use of earphones, use of firearms, etc.
Acute noise-induced damage. The immediate effects of exposure to high-intensity sound stimuli (for example, explosions) include elevation of the hearing threshold, rupture of the eardrum, and traumatic damage to the middle and inner ears (dislocation of ossicles, cochlear injury or fistulas).
Temporary threshold shift. Noise exposure results in a decrease in the sensitivity of auditory sensory cells which is proportional to the duration and intensity of exposure. In its early stages, this increase in auditory threshold, known as auditory fatigue or temporary threshold shift (TTS), is entirely reversible but persists for some time after the cessation of exposure.
Studies of the recovery of auditory sensitivity have identified several types of auditory fatigue. Short-term fatigue dissipates in less than two minutes and results in a maximum threshold shift at the exposure frequency. Long-term fatigue is characterized by recovery in more than two minutes but less than 16 hours, an arbitrary limit derived from studies of industrial noise exposure. In general, auditory fatigue is a function of stimulus intensity, duration, frequency, and continuity. Thus, for a given dose of noise, obtained by integration of intensity and duration, intermittent exposure patterns are less harmful than continuous ones.
The severity of the TTS increases by approximately 6 dB for every doubling of stimulus intensity. Above a specific exposure intensity (the critical level), this rate increases, particularly if exposure is to impulse noise. The TTS increases asymptotically with exposure duration; the asymptote itself increases with stimulus intensity. Due to the characteristics of the outer and middle ears’ transfer function, low frequencies are tolerated the best.
Studies on exposure to pure tones indicate that as the stimulus intensity increases, the frequency at which the TTS is the greatest progressively shifts towards frequencies above that of the stimulus. Subjects exposed to a pure tone of 2,000 Hz develop TTS which is maximal at approximately 3,000 Hz (a shift of a semi-octave). The noise’s effect on the outer hair cells is believed to be responsible for this phenomenon.
The worker who shows TTS recovers to baseline hearing values within hours after removal from noise. However, repeated noise exposures result in less hearing recovery and resultant permanent hearing loss.
Permanent threshold shift. Exposure to high-intensity sound stimuli over several years may lead to permanent hearing loss. This is referred to as permanent threshold shift (PTS). Anatomically, PTS is characterized by degeneration of the hair cells, starting with slight histological modifications but eventually culminating in complete cell destruction. Hearing loss is most likely to involve frequencies to which the ear is most sensitive, as it is at these frequencies that the transmission of acoustic energy from the external environment to the inner ear is optimal. This explains why hearing loss at 4,000 Hz is the first sign of occupationally induced hearing loss (figure 3). Interaction has been observed between stimulus intensity and duration, and international standards assume the degree of hearing loss to a function of the total acoustic energy received by the ear (dose of noise).
Figure 3. Audiogram showing bilateral noise-induced hearing loss.
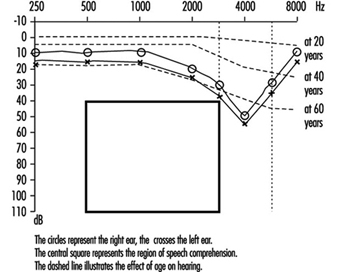
The development of noise-induced hearing loss shows individual susceptibility. Various potentially important variables have been examined to explain this susceptibility, such as age, gender, race, cardiovascular disease, smoking, etc. The data were inconclusive.
An interesting question is whether the amount of TTS could be used to predict the risk of PTS. As noted above, there is a progressive shift of the TTS to frequencies above that of the stimulation frequency. On the other hand, most of the ciliary damage occurring at high stimulus intensities involves cells that are sensitive to the stimulus frequency. Should exposure persist, the difference between the frequency at which the PTS is maximal and the stimulation frequency progressively decreases. Ciliary damage and cell loss consequently occurs in the cells most sensitive to the stimulus frequencies. It thus appears that TTS and PTS involve different mechanisms, and that it is thus impossible to predict an individual’s PTS on the basis of the observed TTS.
Individuals with PTS are usually asymptomatic initially. As the hearing loss progresses, they begin to have difficulty following conversations in noisy settings such as parties or restaurants. The progression, which usually affects the ability to perceive high-pitched sounds first, is usually painless and relatively slow.
Examination of individuals suffering from hearing loss
Clinical examination
In addition to the history of the date when the hearing loss was first detected (if any) and how it has evolved, including any asymmetry of hearing, the medical questionnaire should elicit information on the patient’s age, family history, use of ototoxic medications or exposure to other ototoxic chemicals, the presence of tinnitus (i.e., buzzing, whistling or ringing sounds in one or both ears), dizziness or any problems with balance, and any history of ear infections with pain or discharge from the outer ear canal. Of critical importance is a detailed life-long history of exposures to high sound levels (note that, to the layperson, not all sounds are “noise”) on the job, in previous jobs and off-the-job. A history of episodes of TTS would confirm prior toxic exposures to noise.
Physical examination should include evaluation of the function of the other cranial nerves, tests of balance, and ophthalmoscopy to detect any evidence of increased cranial pressure. Visual examination of the external auditory canal will detect any impacted cerumen and, after it has been cautiously removed (no sharp object!), any evidence of scarring or perforation of the tympanic membrane. Hearing loss can be determined very crudely by testing the patient’s ability to repeat words and phrases spoken softly or whispered by the examiner when positioned behind and out of the sight of the patient. The Weber test (placing a vibrating tuning fork in the centre of the forehead to determine if this sound is “heard” in either or both ears) and the Rinné pitch-pipe test (placing a vibrating tuning fork on the mastoid process until the patient can no longer hear the sound, then quickly placing the fork near the ear canal; normally the sound can be heard longer through air than through bone) will allow classification of the hearing loss as transmission- or neurosensory.
The audiogram is the standard test to detect and evaluate hearing loss (see below). Specialized studies to complement the audiogram may be necessary in some patients. These include: tympanometry, word discrimination tests, evaluation of the attenuation reflex, electrophysical studies (electrocochleogram, auditory evoked potentials) and radiological studies (routine skull x rays complemented by CAT scan, MRI).
Audiometry
This crucial component of the medical evaluation uses a device known as an audiometer to determine the auditory threshold of individuals to pure tones of 250-8,000 Hz and sound levels between –10 dB (the hearing threshold of intact ears) and 110 dB (maximal damage). To eliminate the effects of TTSs, patients should not have been exposed to noise during the previous 16 hours. Air conduction is measured by earphones placed on the ears, while bone conduction is measured by placing a vibrator in contact with the skull behind the ear. Each ear’s hearing is measured separately and test results are reported on a graph known as an audiogram (Figure 3). The threshold of intelligibility, that is. the sound intensity at which speech becomes intelligible, is determined by a complementary test method known as vocal audiometry, based on the ability to understand words composed of two syllables of equal intensity (for instance, shepherd, dinner, stunning).
Comparison of air and bone conduction allows classification of hearing losses as transmission (involving the external auditory canal or middle ear) or neurosensory loss (involving the inner ear or auditory nerve) (figures 3 and 4). The audiogram observed in cases of noise-induced hearing loss is characterized by an onset of hearing loss at 4,000 Hz, visible as a dip in the audiogram (figure 3). As exposure to excessive noise levels continues, neighbouring frequencies are progressively affected and the dip broadens, encroaching, at approximately 3,000 Hz, on frequencies essential for the comprehension of conversation. Noise-induced hearing loss is usually bilateral and shows a similar pattern in both ears, that is, the difference between the two ears does not exceed 15 dB at 500 Hz, at 1,000 dB and at 2,000 Hz, and 30 dB at 3,000, at 4,000 and at 6,000 Hz. Asymmetric damage may, however, be present in cases of non-uniform exposure, for example, with marksmen, in whom hearing loss is higher on the side opposite to the trigger finger (the left side, in a right-handed person). In hearing loss unrelated to noise exposure, the audiogram does not exhibit the characteristic 4,000 Hz dip (figure 4).
Figure 4. Examples of right-ear audiograms. The circles represent air-conduction hearing loss, the ““ bone conduction.
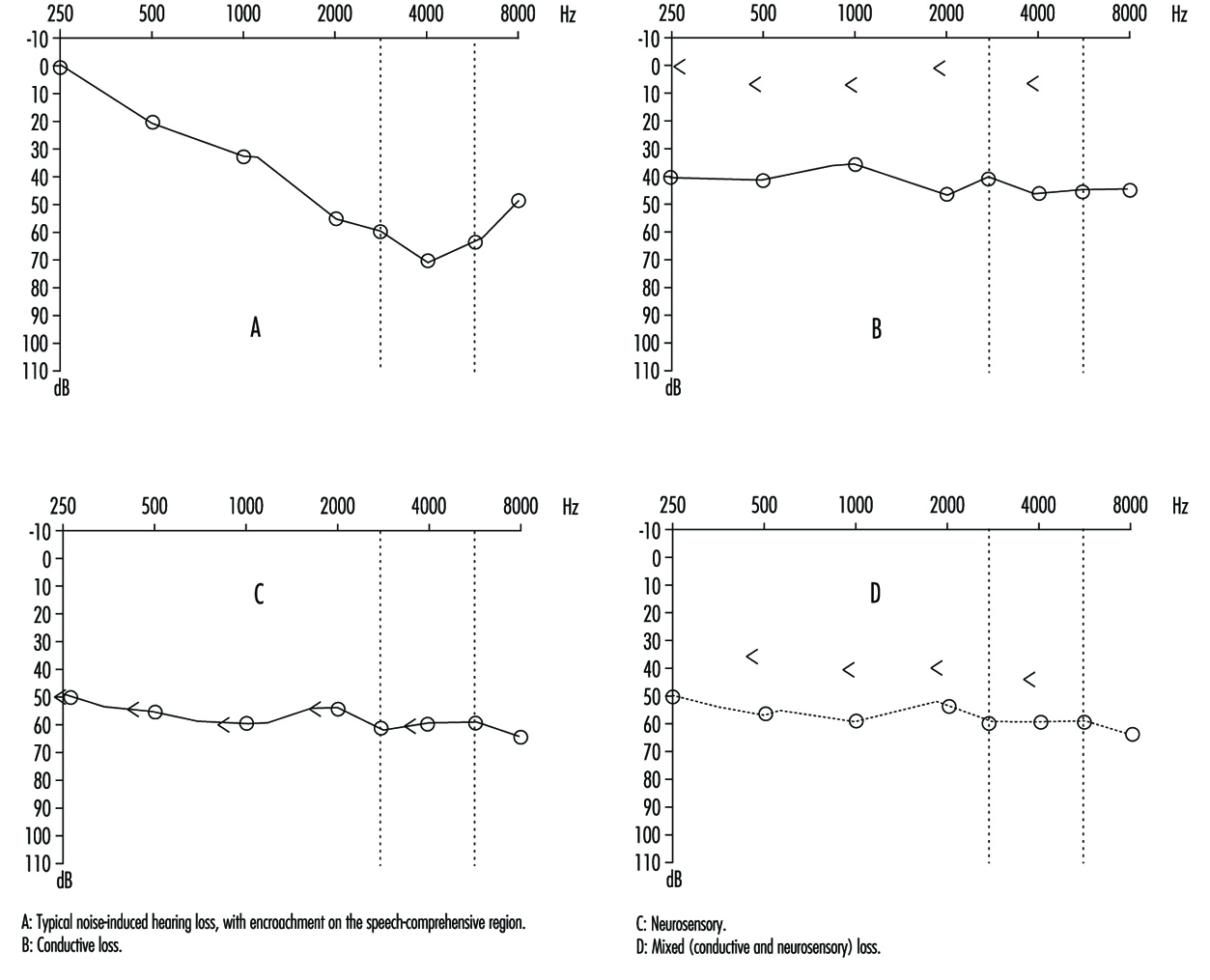
There are two types of audiometric examinations: screening and diagnostic. Screening audiometry is used for the rapid examination of groups of individuals in the workplace, in schools or elsewhere in the community to identify those who appear to have some hearing loss. Often, electronic audiometers that permit self-testing are used and, as a rule, screening audiograms are obtained in a quiet area but not necessarily in a sound-proof, vibration-free chamber. The latter is considered to be a prerequisite for diagnostic audiometry which is intended to measure hearing loss with reproducible precision and accuracy. The diagnostic examination is properly performed by a trained audiologist (in some circumstances, formal certification of the competence of the audiologist is required). The accuracy of both types of audiometry depends on periodic testing and recalibration of the equipment being used.
In many jurisdictions, individuals with job-related, noise-induced hearing loss are eligible for workers’ compensation benefits. Accordingly, many employers are including audiometry in their preplacement medical examinations to detect any existing hearing loss that may be the responsibility of a previous employer or represent a non-occupational exposure.
Hearing thresholds progressively increase with age, with higher frequencies being more affected (figure 3). The characteristic 4,000 Hz dip observed in noise-induced hearing loss is not seen with this type of hearing loss.
Calculation of hearing loss
In the United States the most widely accepted formula for calculating functional limitation related to hearing loss is the one proposed in 1979 by the American Academy of Otolaryngology (AAO) and adopted by the American Medical Association. It is based on the average of values obtained at 500, at 1,000, at 2,000 and at 3,000 Hz (table 1), with the lower limit for functional limitation set at 25 dB.
Table 1. Typical calculation of functional loss from an audiogram
| Frequency | |||||||
| 500 Hz |
1,000 Hz |
2,000 Hz |
3,000 Hz |
4,000 Hz |
6,000 Hz |
8,000 Hz |
|
| Right ear (dB) | 25 | 35 | 35 | 45 | 50 | 60 | 45 |
| Left ear (dB) | 25 | 35 | 40 | 50 | 60 | 70 | 50 |
| Unilateral loss |
| Percentage unilateral loss = (average at 500, 1,000, 2,000 and 3,000 Hz) – 25dB (lower limit) x1.5 |
| Example: Right ear: [([25 + 35 + 35 + 45]/4) – 25) x 1.5 = 15 (per cent) Left ear: [([25 + 35 + 40 + 50]/4) – 25) x 1.5 = 18.8 (per cent) |
| Bilateral loss |
| Percentage of bilateral loss = {(percentage of unilateral loss of the best ear x 5) + (percentage of unilateral loss of the worst ear)}/6 |
| Example: {(15 x 5) + 18.8}/6 = 15.6 (per cent) |
Source: Rees and Duckert 1994.
Presbycusis
Presbycusis or age-related hearing loss generally begins at about age 40 and progresses gradually with increasing age. It is usually bilateral. The characteristic 4,000 Hz dip observed in noise-induced hearing loss is not seen with presbycusis. However, it is possible to have the effects of ageing superimposed on noise-related hearing loss.
Treatment
The first essential of treatment is avoidance of any further exposure to potentially toxic levels of noise (see “Prevention” below). It is generally believed that no more subsequent hearing loss occurs after the removal from noise exposure than would be expected from the normal ageing process.
While conduction losses, for example, those related to acute traumatic noise-induced damage, are amenable to medical treatment or surgery, chronic noise-induced hearing loss cannot be corrected by treatment. The use of a hearing aid is the sole “remedy” possible, and is only indicated when hearing loss affects the frequencies critical for speech comprehension (500 to 3,000 Hz). Other types of support, for example lip-reading and sound amplifiers (on telephones, for example), may, however, be possible.
Prevention
Because noise-induced hearing loss is permanent, it is essential to apply any measure likely to reduce exposure. This includes reduction at the source (quieter machines and equipment or encasing them in sound-proof enclosures) or the use of individual protective devices such as ear plugs and/or ear muffs. If reliance is placed on the latter, it is imperative to verify that their manufacturers’ claims for effectiveness are valid and that exposed workers are using them properly at all times.
The designation of 85 dB (A) as the highest permissible occupational exposure limit was to protect the greatest number of people. But, since there is significant interpersonal variation, strenuous efforts to keep exposures well below that level are indicated. Periodic audiometry should be instituted as part of the medical surveillance programme to detect as early as possible any effects that may indicate noise toxicity.