- Vous êtes ici:
-
Accueil

-
Table des matières
-
XVIIe partie. Services et Commerce
-
Divertissement et arts
- Arts et Métiers
Toxicologie dans la réglementation de la santé et de la sécurité
La toxicologie joue un rôle majeur dans l'élaboration des réglementations et autres politiques de santé au travail. Afin de prévenir les accidents du travail et les maladies professionnelles, les décisions sont de plus en plus fondées sur des informations pouvant être obtenues avant ou en l'absence des types d'expositions humaines qui fourniraient des informations définitives sur les risques, telles que des études épidémiologiques. De plus, les études toxicologiques, telles que décrites dans ce chapitre, peuvent fournir des informations précises sur la dose et la réponse dans les conditions contrôlées de la recherche en laboratoire ; ces informations sont souvent difficiles à obtenir dans le cadre d'expositions professionnelles non contrôlées. Cependant, ces informations doivent être soigneusement évaluées afin d'estimer la probabilité d'effets nocifs chez l'homme, la nature de ces effets nocifs et la relation quantitative entre les expositions et les effets.
Une attention considérable a été accordée dans de nombreux pays, depuis les années 1980, à l'élaboration de méthodes objectives d'utilisation des informations toxicologiques dans la prise de décision réglementaire. Les méthodes formelles, souvent appelées évaluation des risques, ont été proposés et utilisés dans ces pays par des entités gouvernementales et non gouvernementales. L'évaluation des risques a été diversement définie; fondamentalement, il s'agit d'un processus d'évaluation qui intègre des informations sur la toxicologie, l'épidémiologie et l'exposition pour identifier et estimer la probabilité d'effets indésirables associés à l'exposition à des substances ou conditions dangereuses. L'évaluation des risques peut être de nature qualitative, indiquant la nature d'un effet nocif et une estimation générale de la probabilité, ou elle peut être quantitative, avec des estimations du nombre de personnes affectées à des niveaux d'exposition spécifiques. Dans de nombreux systèmes réglementaires, l'évaluation des risques se déroule en quatre étapes : identification des dangers, la description de la nature de l'effet toxique ; évaluation dose-réponse, une analyse semi-quantitative ou quantitative de la relation entre l'exposition (ou la dose) et la gravité ou la probabilité de l'effet toxique ; évaluation de l'exposition, l'évaluation des informations sur la gamme d'expositions susceptibles de se produire pour les populations en général ou pour des sous-groupes au sein des populations ; caractérisation des risques, la compilation de toutes les informations ci-dessus dans une expression de l'ampleur du risque susceptible de se produire dans des conditions d'exposition spécifiées (voir CNRC 1983 pour un énoncé de ces principes).
Dans cette section, trois approches d'évaluation des risques sont présentées à titre d'illustration. Il est impossible de fournir un recueil complet des méthodes d'évaluation des risques utilisées dans le monde, et ces sélections ne doivent pas être considérées comme prescriptives. Il convient de noter qu'il existe une tendance à l'harmonisation des méthodes d'évaluation des risques, en partie en réponse aux dispositions des récents accords du GATT. Deux processus d'harmonisation internationale des méthodes d'évaluation des risques sont actuellement en cours, à travers le Programme international sur la sécurité chimique (IPCS) et l'Organisation de coopération et de développement économiques (OCDE). Ces organisations tiennent également à jour des informations sur les approches nationales de l'évaluation des risques.
Structurer les relations d'activité
L'analyse des relations structure-activité (SAR) consiste à utiliser des informations sur la structure moléculaire des produits chimiques pour prédire des caractéristiques importantes liées à la persistance, à la distribution, à l'absorption et à l'absorption et à la toxicité. Le SAR est une méthode alternative d'identification des produits chimiques potentiellement dangereux, qui promet d'aider les industries et les gouvernements à hiérarchiser les substances pour une évaluation plus approfondie ou pour la prise de décision à un stade précoce pour de nouveaux produits chimiques. La toxicologie est une entreprise de plus en plus coûteuse et gourmande en ressources. Les préoccupations croissantes concernant le potentiel des produits chimiques à causer des effets néfastes sur les populations humaines exposées ont incité les organismes de réglementation et de santé à élargir la gamme et la sensibilité des tests pour détecter les risques toxicologiques. Dans le même temps, les charges réelles et perçues de la réglementation sur l'industrie ont suscité des inquiétudes quant à l'aspect pratique des méthodes d'essais de toxicité et de l'analyse des données. À l'heure actuelle, la détermination de la cancérogénicité chimique dépend de tests sur la durée de vie d'au moins deux espèces, des deux sexes, à plusieurs doses, avec une analyse histopathologique minutieuse de plusieurs organes, ainsi que la détection de changements prénéoplasiques dans les cellules et les organes cibles. Aux États-Unis, on estime que le test biologique du cancer coûte plus de 3 millions de dollars (dollars de 1995).
Même avec des ressources financières illimitées, la charge de tester les quelque 70,000 1984 produits chimiques existants produits dans le monde aujourd'hui dépasserait les ressources disponibles des toxicologues qualifiés. Des siècles seraient nécessaires pour réaliser ne serait-ce qu'une évaluation de premier niveau de ces produits chimiques (NRC 1993). Dans de nombreux pays, les préoccupations éthiques concernant l'utilisation d'animaux dans les tests de toxicité ont augmenté, ce qui exerce des pressions supplémentaires sur l'utilisation des méthodes standard de test de toxicité. Le SAR a été largement utilisé dans l'industrie pharmaceutique pour identifier les molécules ayant un potentiel d'utilisation bénéfique dans le traitement (Hansch et Zhang 1979). Dans la politique de santé environnementale et professionnelle, le SAR est utilisé pour prédire la dispersion des composés dans l'environnement physico-chimique et pour sélectionner de nouveaux produits chimiques pour une évaluation plus approfondie de la toxicité potentielle. En vertu de la Toxic Substances Control Act (TSCA) des États-Unis, l'EPA utilise depuis 5 une approche SAR comme « premier crible » des nouveaux produits chimiques dans le processus de notification avant fabrication (PMN) ; L'Australie utilise une approche similaire dans le cadre de sa procédure de notification des nouveaux produits chimiques (NICNAS). Aux États-Unis, l'analyse SAR est une base importante pour déterminer qu'il existe une base raisonnable pour conclure que la fabrication, le traitement, la distribution, l'utilisation ou l'élimination de la substance présentera un risque déraisonnable de préjudice pour la santé humaine ou l'environnement, comme l'exige la section 6(f) de la TSCA. Sur la base de cette découverte, l'EPA peut alors exiger des tests réels de la substance en vertu de la section XNUMX de la TSCA.
Justification du SAR
La justification scientifique du SAR est basée sur l'hypothèse que la structure moléculaire d'un produit chimique prédira des aspects importants de son comportement dans les systèmes physico-chimiques et biologiques (Hansch et Leo 1979).
Processus SAR
Le processus d'examen SAR comprend l'identification de la structure chimique, y compris les formulations empiriques ainsi que le composé pur ; identification de substances structurellement analogues ; rechercher des bases de données et de la littérature pour obtenir des informations sur les analogues structuraux ; et l'analyse de la toxicité et d'autres données sur les analogues structuraux. Dans de rares cas, les informations sur la structure du composé à elles seules peuvent être suffisantes pour étayer certaines analyses SAR, basées sur des mécanismes de toxicité bien compris. Plusieurs bases de données sur le SAR ont été compilées, ainsi que des méthodes informatiques pour la prédiction de la structure moléculaire.
Avec ces informations, les paramètres suivants peuvent être estimés avec SAR :
- paramètres physico-chimiques : point d'ébullition, pression de vapeur, solubilité dans l'eau, coefficient de partage octanol/eau
- paramètres du devenir biologique/environnemental : biodégradation, sorption dans le sol, photodégradation, pharmacocinétique
- paramètres de toxicité : toxicité pour les organismes aquatiques, absorption, toxicité aiguë pour les mammifères (test limite ou LD50), irritation cutanée, pulmonaire et oculaire, sensibilisation, toxicité subchronique, mutagénicité.
Il convient de noter qu'il n'existe pas de méthodes SAR pour des paramètres de santé aussi importants que la cancérogénicité, la toxicité pour le développement, la toxicité pour la reproduction, la neurotoxicité, l'immunotoxicité ou d'autres effets sur les organes cibles. Cela est dû à trois facteurs : l'absence d'une grande base de données sur laquelle tester les hypothèses SAR, le manque de connaissances sur les déterminants structurels de l'action toxique et la multiplicité des cellules cibles et des mécanismes impliqués dans ces paramètres (voir "The United States approche d'évaluation des risques des substances toxiques pour la reproduction et des agents neurotoxiques »). Quelques tentatives limitées d'utilisation du SAR pour prédire la pharmacocinétique en utilisant des informations sur les coefficients de partage et la solubilité (Johanson et Naslund 1988). Un SAR quantitatif plus étendu a été réalisé pour prédire le métabolisme dépendant du P450 d'une gamme de composés et la liaison des molécules de type dioxine et PCB au récepteur cytosolique de la « dioxine » (Hansch et Zhang 1993).
Il a été démontré que le DAS a une prévisibilité variable pour certains des paramètres énumérés ci-dessus, comme indiqué dans le tableau 1. Ce tableau présente les données de deux comparaisons de l'activité prévue avec les résultats réels obtenus par des mesures empiriques ou des tests de toxicité. Le SAR, tel qu'il a été mené par des experts de l'US EPA, a obtenu de moins bons résultats pour prédire les propriétés physico-chimiques que pour prédire l'activité biologique, y compris la biodégradation. Pour les paramètres de toxicité, le SAR a obtenu les meilleurs résultats pour prédire la mutagénicité. Ashby et Tennant (1991) dans une étude plus approfondie ont également trouvé une bonne prévisibilité de la génotoxicité à court terme dans leur analyse des produits chimiques NTP. Ces résultats ne sont pas surprenants, compte tenu des connaissances actuelles sur les mécanismes moléculaires de la génotoxicité (voir « Toxicologie génétique ») et le rôle de l'électrophilie dans la liaison à l'ADN. En revanche, le SAR avait tendance à sous-estimer la toxicité systémique et subchronique chez les mammifères et à surestimer la toxicité aiguë pour les organismes aquatiques.
Tableau 1. Comparaison des données SAR et des tests : analyses OCDE/NTP
| Endpoint | Une entente (%) | Désaccord (%) | Numéro |
| Point d'ébullition | 50 | 50 | 30 |
| Pression de vapeur | 63 | 37 | 113 |
| Solubilité dans l'eau | 68 | 32 | 133 |
| Coefficient de partage | 61 | 39 | 82 |
| Biodégradation | 93 | 7 | 107 |
| Toxicité pour les poissons | 77 | 22 | 130 |
| Toxicité de la daphnie | 67 | 33 | 127 |
| Toxicité aiguë pour les mammifères (DL50 ) | 80 | 201 | 142 |
| Irritation de la peau | 82 | 18 | 144 |
| Irritation de l'oeil | 78 | 22 | 144 |
| Sensibilisation cutanée | 84 | 16 | 144 |
| Toxicité subchronique | 57 | 32 | 143 |
| Mutagénicité2 | 88 | 12 | 139 |
| Mutagénicité3 | 82-944 | 1-10 | 301 |
| Cancérogénicité3 : Essai biologique de deux ans | 72-954 | - | 301 |
Source : Données de l'OCDE, communication personnelle C. Auer, US EPA. Seuls les paramètres pour lesquels des prédictions de DAS comparables et des données de test réelles étaient disponibles ont été utilisés dans cette analyse. Les données du NTP proviennent d'Ashby et Tennant 1991.
1 L'incapacité du SAR à prédire la toxicité aiguë de 12 % des produits chimiques testés était préoccupante.
2 Données OCDE, basées sur la concordance du test d'Ames avec le DAS
3 Données du NTP, basées sur des analyses de genetox comparées aux prévisions de DAS pour plusieurs classes de « produits chimiques structurellement alertants ».
4 La concordance varie selon la classe; la concordance la plus élevée était avec les composés amino/nitro aromatiques; le plus bas avec des structures « diverses ».
Pour d'autres paramètres toxiques, comme indiqué ci-dessus, le DAS a une utilité moins démontrable. Les prévisions de toxicité pour les mammifères sont compliquées par le manque de SAR pour la toxicocinétique des molécules complexes. Néanmoins, certaines tentatives ont été faites pour proposer des principes SAR pour des critères complexes de toxicité pour les mammifères (par exemple, voir Bernstein (1984) pour une analyse SAR des substances potentiellement toxiques pour la reproduction mâle). Dans la plupart des cas, la base de données est trop petite pour permettre des tests rigoureux des prédictions basées sur la structure.
À ce stade, on peut conclure que le SAR peut être utile principalement pour hiérarchiser l'investissement dans les ressources d'essais de toxicité ou pour soulever rapidement des préoccupations concernant un danger potentiel. Ce n'est qu'en cas de mutagénicité qu'il est probable que l'analyse SAR en elle-même puisse être utilisée avec fiabilité pour éclairer d'autres décisions. Pour aucun effet, il est probable que le DAS puisse fournir le type d'informations quantitatives requises à des fins d'évaluation des risques, comme indiqué ailleurs dans ce chapitre et Encyclopédie.
Tests de toxicité in vitro
L'émergence de technologies sophistiquées en biologie moléculaire et cellulaire a stimulé une évolution relativement rapide dans les sciences de la vie, y compris la toxicologie. En effet, le centre d'intérêt de la toxicologie se déplace des animaux entiers et des populations d'animaux entiers vers les cellules et les molécules d'animaux et d'humains individuels. Depuis le milieu des années 1980, les toxicologues ont commencé à utiliser ces nouvelles méthodologies pour évaluer les effets des produits chimiques sur les systèmes vivants. En tant que progression logique, ces méthodes sont adaptées aux fins d'essais de toxicité. Ces avancées scientifiques se sont conjuguées à des facteurs sociaux et économiques pour modifier l'évaluation de la sécurité des produits et des risques potentiels.
Les facteurs économiques sont spécifiquement liés au volume de matériaux qui doivent être testés. Une multitude de nouveaux produits cosmétiques, pharmaceutiques, pesticides, chimiques et ménagers sont introduits sur le marché chaque année. Tous ces produits doivent être évalués pour leur toxicité potentielle. En outre, il existe un arriéré de produits chimiques déjà utilisés qui n'ont pas été suffisamment testés. L'énorme tâche d'obtenir des informations détaillées sur la sécurité de tous ces produits chimiques en utilisant des méthodes traditionnelles d'expérimentation sur des animaux entiers serait coûteuse en termes d'argent et de temps, si elle pouvait même être accomplie.
Il existe également des problèmes de société liés à la santé et à la sécurité publiques, ainsi qu'une préoccupation croissante du public concernant l'utilisation d'animaux pour les tests de sécurité des produits. En ce qui concerne la sécurité humaine, les groupes d'intérêt public et de défense de l'environnement ont exercé une pression importante sur les agences gouvernementales pour qu'elles appliquent des réglementations plus strictes sur les produits chimiques. Un exemple récent de cela a été un mouvement de certains groupes environnementaux pour interdire le chlore et les composés contenant du chlore aux États-Unis. L'une des motivations d'une telle action extrême réside dans le fait que la plupart de ces composés n'ont jamais été suffisamment testés. D'un point de vue toxicologique, le concept d'interdire toute une classe de produits chimiques divers sur la simple base de la présence de chlore est à la fois scientifiquement non fondé et irresponsable. Pourtant, il est compréhensible que du point de vue du public, il doit y avoir une certaine assurance que les produits chimiques rejetés dans l'environnement ne posent pas de risque important pour la santé. Une telle situation souligne la nécessité de méthodes plus efficaces et rapides pour évaluer la toxicité.
L'autre préoccupation sociétale qui a eu un impact sur le domaine des tests de toxicité est le bien-être animal. Le nombre croissant de groupes de protection des animaux à travers le monde ont exprimé une opposition considérable à l'utilisation d'animaux entiers pour les tests de sécurité des produits. Des campagnes actives ont été menées contre les fabricants de cosmétiques, de produits ménagers et de soins personnels et de produits pharmaceutiques pour tenter d'arrêter les tests sur les animaux. Ces efforts en Europe ont abouti à l'adoption du sixième amendement à la directive 76/768/CEE (la directive sur les cosmétiques). La conséquence de cette directive est que les produits cosmétiques ou les ingrédients cosmétiques qui ont été testés sur des animaux après le 1er janvier 1998 ne peuvent être commercialisés dans l'Union européenne, sauf si des méthodes alternatives sont insuffisamment validées. Bien que cette directive n'ait aucune compétence sur la vente de ces produits aux États-Unis ou dans d'autres pays, elle affectera de manière significative les entreprises qui ont des marchés internationaux qui incluent l'Europe.
La notion d'alternatives, qui est à la base du développement de tests autres que ceux sur animaux entiers, est définie par les trois Rs: réduction du nombre d'animaux utilisés; raffinement de protocoles pour que les animaux ressentent moins de stress ou d'inconfort ; et remplacement des tests actuels sur les animaux avec des tests in vitro (c'est-à-dire des tests effectués en dehors de l'animal vivant), des modèles informatiques ou des tests sur des vertébrés inférieurs ou des espèces d'invertébrés. Les trois Rs ont été introduits dans un livre publié en 1959 par deux scientifiques britanniques, WMS Russell et Rex Burch, Les principes de la technique expérimentale humaine. Russell et Burch ont soutenu que la seule façon d'obtenir des résultats scientifiques valables était le traitement humain des animaux et pensaient que des méthodes devraient être développées pour réduire l'utilisation des animaux et finalement la remplacer. Fait intéressant, les principes énoncés par Russell et Burch ont reçu peu d'attention jusqu'à la résurgence du mouvement de protection des animaux au milieu des années 1970. Aujourd'hui, le concept des trois Rs est très à l'avant-garde en matière de recherche, d'essais et d'éducation.
En résumé, le développement de méthodologies d'essais in vitro a été influencé par une variété de facteurs qui ont convergé au cours des dix à vingt dernières années. Il est difficile de déterminer si l'un de ces facteurs à lui seul aurait eu un effet aussi profond sur les stratégies d'essais de toxicité.
Concept des tests de toxicité in vitro
Cette section se concentrera uniquement sur les méthodes in vitro d'évaluation de la toxicité, comme l'une des alternatives aux tests sur l'animal entier. D'autres alternatives non animales telles que la modélisation informatique et les relations quantitatives structure-activité sont abordées dans d'autres articles de ce chapitre.
Les études in vitro sont généralement menées sur des cellules ou des tissus animaux ou humains à l'extérieur du corps. In vitro signifie littéralement « dans du verre », et fait référence à des procédures effectuées sur du matériel vivant ou des composants de matériel vivant cultivés dans des boîtes de Pétri ou dans des tubes à essai dans des conditions définies. Celles-ci peuvent être opposées aux études in vivo, ou celles réalisées « chez l'animal vivant ». Bien qu'il soit difficile, voire impossible, de projeter les effets d'un produit chimique sur un organisme complexe lorsque les observations se limitent à un seul type de cellules dans une boîte, les études in vitro fournissent également une quantité importante d'informations sur la toxicité intrinsèque. que les mécanismes cellulaires et moléculaires de la toxicité. De plus, elles offrent de nombreux avantages par rapport aux études in vivo en ce sens qu'elles sont généralement moins coûteuses et qu'elles peuvent être réalisées dans des conditions plus contrôlées. De plus, malgré le fait qu'un petit nombre d'animaux sont encore nécessaires pour obtenir des cellules pour les cultures in vitro, ces méthodes peuvent être considérées comme des alternatives de réduction (puisque beaucoup moins d'animaux sont utilisés par rapport aux études in vivo) et des alternatives de raffinement (car elles éliminent le besoin soumettre les animaux aux conséquences toxiques néfastes imposées par les expériences in vivo).
Afin d'interpréter les résultats des tests de toxicité in vitro, de déterminer leur utilité potentielle dans l'évaluation de la toxicité et de les relier au processus toxicologique global in vivo, il est nécessaire de comprendre quelle partie du processus toxicologique est examinée. L'ensemble du processus toxicologique consiste en des événements qui commencent par l'exposition de l'organisme à un agent physique ou chimique, progressent par des interactions cellulaires et moléculaires et se manifestent finalement dans la réponse de l'organisme entier. Les tests in vitro sont généralement limités à la partie du processus toxicologique qui se déroule au niveau cellulaire et moléculaire. Les types d'informations pouvant être obtenues à partir d'études in vitro comprennent les voies métaboliques, l'interaction des métabolites actifs avec des cibles cellulaires et moléculaires et des paramètres toxiques potentiellement mesurables qui peuvent servir de biomarqueurs moléculaires pour l'exposition. Dans une situation idéale, le mécanisme de toxicité de chaque produit chimique résultant de l'exposition à la manifestation de l'organisme serait connu, de sorte que les informations obtenues à partir des tests in vitro pourraient être entièrement interprétées et liées à la réponse de l'organisme entier. Cependant, cela est pratiquement impossible, puisque relativement peu de mécanismes toxicologiques complets ont été élucidés. Ainsi, les toxicologues sont confrontés à une situation dans laquelle les résultats d'un test in vitro ne peuvent pas être utilisés comme une prédiction entièrement précise de la toxicité in vivo car le mécanisme est inconnu. Cependant, fréquemment au cours du processus de développement d'un test in vitro, les composants du ou des mécanismes cellulaires et moléculaires de la toxicité sont élucidés.
L'une des principales questions non résolues entourant le développement et la mise en œuvre des tests in vitro est liée à la considération suivante : doivent-ils être basés sur des mécanismes ou suffit-il qu'ils soient descriptifs ? Il est incontestablement préférable, d'un point de vue scientifique, de n'utiliser que des tests basés sur des mécanismes pour remplacer les tests in vivo. Cependant, en l'absence de connaissances mécanistes complètes, la perspective de développer des tests in vitro pour remplacer complètement les tests sur des animaux entiers dans un avenir proche est presque nulle. Cela n'exclut toutefois pas l'utilisation de types de tests plus descriptifs comme outils de dépistage précoce, ce qui est le cas actuellement. Ces écrans ont entraîné une réduction significative de l'utilisation des animaux. Par conséquent, jusqu'à ce que des informations plus mécanistes soient générées, il peut être nécessaire d'employer, dans une mesure plus limitée, des tests dont les résultats sont simplement bien corrélés avec ceux obtenus in vivo.
Tests in vitro de cytotoxicité
Dans cette section, plusieurs tests in vitro qui ont été développés pour évaluer le potentiel cytotoxique d'un produit chimique seront décrits. Pour la plupart, ces tests sont faciles à réaliser et l'analyse peut être automatisée. Un test in vitro couramment utilisé pour la cytotoxicité est le test au rouge neutre. Ce test est effectué sur des cellules en culture et, pour la plupart des applications, les cellules peuvent être maintenues dans des boîtes de culture contenant 96 petits puits de 6.4 mm de diamètre chacun. Étant donné que chaque puits peut être utilisé pour une seule détermination, cet arrangement peut accueillir plusieurs concentrations du produit chimique d'essai ainsi que des contrôles positifs et négatifs avec un nombre suffisant de répétitions pour chacun. Après traitement des cellules avec diverses concentrations du produit chimique d'essai allant d'au moins deux ordres de grandeur (par exemple, de 0.01 mM à 1 mM), ainsi que des produits chimiques témoins positifs et négatifs, les cellules sont rincées et traitées avec du rouge neutre, un colorant qui ne peut être absorbé et retenu que par les cellules vivantes. Le colorant peut être ajouté lors du retrait du produit chimique d'essai pour déterminer les effets immédiats, ou il peut être ajouté à différents moments après le retrait du produit chimique d'essai pour déterminer les effets cumulatifs ou différés. L'intensité de la couleur dans chaque puits correspond au nombre de cellules vivantes dans ce puits. L'intensité de la couleur est mesurée par un spectrophotomètre qui peut être équipé d'un lecteur de plaques. Le lecteur de plaque est programmé pour fournir des mesures individuelles pour chacun des 96 puits de la boîte de culture. Cette méthodologie automatisée permet à l'investigateur d'effectuer rapidement une expérience concentration-réponse et d'obtenir des données statistiquement utiles.
Un autre test relativement simple de cytotoxicité est le test MTT. Le MTT (bromure de 3[4,5-diméthylthiazol-2-yl]-2,5-diphényltétrazolium) est un colorant tétrazolium qui est réduit par les enzymes mitochondriales à une couleur bleue. Seules les cellules possédant des mitochondries viables conserveront la capacité de réaliser cette réaction ; par conséquent, l'intensité de la couleur est directement liée au degré d'intégrité mitochondriale. Il s'agit d'un test utile pour détecter les composés cytotoxiques généraux ainsi que les agents qui ciblent spécifiquement les mitochondries.
La mesure de l'activité de la lactate déshydrogénase (LDH) est également utilisée comme test à grande échelle pour la cytotoxicité. Cette enzyme est normalement présente dans le cytoplasme des cellules vivantes et est libérée dans le milieu de culture cellulaire par les membranes cellulaires non étanches des cellules mortes ou mourantes qui ont été affectées par un agent toxique. De petites quantités de milieu de culture peuvent être retirées à divers moments après le traitement chimique des cellules pour mesurer la quantité de LDH libérée et déterminer une évolution temporelle de la toxicité. Bien que le test de libération de LDH soit une évaluation très générale de la cytotoxicité, il est utile car il est facile à réaliser et peut être effectué en temps réel.
De nombreuses nouvelles méthodes sont en cours de développement pour détecter les dommages cellulaires. Des méthodes plus sophistiquées utilisent des sondes fluorescentes pour mesurer une variété de paramètres intracellulaires, tels que la libération de calcium et les changements de pH et de potentiel de membrane. En général, ces sondes sont très sensibles et peuvent détecter des changements cellulaires plus subtils, réduisant ainsi la nécessité d'utiliser la mort cellulaire comme point final. De plus, bon nombre de ces tests fluorescents peuvent être automatisés par l'utilisation de plaques à 96 puits et de lecteurs de plaques fluorescentes.
Une fois que des données ont été recueillies sur une série de produits chimiques à l'aide de l'un de ces tests, les toxicités relatives peuvent être déterminées. La toxicité relative d'un produit chimique, telle que déterminée dans un essai in vitro, peut être exprimée comme la concentration qui exerce un effet de 50 % sur la réponse finale des cellules non traitées. Cette détermination est appelée CE50 (Effectif Conconcentration pour 50% des cellules) et peut être utilisé pour comparer les toxicités de différents produits chimiques in vitro. (Un terme similaire utilisé pour évaluer la toxicité relative est IC50, indiquant la concentration d'un produit chimique qui provoque une inhibition de 50% d'un processus cellulaire, par exemple, la capacité d'absorber le rouge neutre.) Il n'est pas facile d'évaluer si la toxicité relative in vitro des produits chimiques est comparable à leur relative dans toxicités in vivo, car il existe de nombreux facteurs de confusion dans le système in vivo, tels que la toxicocinétique, le métabolisme, les mécanismes de réparation et de défense. De plus, étant donné que la plupart de ces tests mesurent les paramètres généraux de cytotoxicité, ils ne sont pas basés sur des mécanismes. Par conséquent, l'accord entre les toxicités relatives in vitro et in vivo est simplement corrélatif. Malgré les nombreuses complexités et difficultés d'extrapolation d'in vitro à in vivo, ces tests in vitro s'avèrent très précieux car ils sont simples et peu coûteux à réaliser et peuvent être utilisés comme écrans pour signaler des médicaments ou des produits chimiques hautement toxiques aux premiers stades de la développement.
Toxicité pour les organes cibles
Des tests in vitro peuvent également être utilisés pour évaluer la toxicité spécifique d'un organe cible. Il existe un certain nombre de difficultés associées à la conception de tels tests, la plus notable étant l'incapacité des systèmes in vitro à conserver de nombreuses caractéristiques de l'organe in vivo. Fréquemment, lorsque des cellules sont prélevées sur des animaux et placées en culture, elles ont tendance soit à dégénérer rapidement et/ou à se dédifférencier, c'est-à-dire à perdre leurs fonctions d'organe et à devenir plus génériques. Cela pose un problème en ce que dans un court laps de temps, généralement quelques jours, les cultures ne sont plus utiles pour évaluer les effets spécifiques d'un organe d'une toxine.
Beaucoup de ces problèmes sont en train d'être surmontés grâce aux progrès récents de la biologie moléculaire et cellulaire. Les informations obtenues sur l'environnement cellulaire in vivo peuvent être utilisées pour moduler les conditions de culture in vitro. Depuis le milieu des années 1980, de nouveaux facteurs de croissance et cytokines ont été découverts, et nombre d'entre eux sont maintenant disponibles dans le commerce. L'ajout de ces facteurs aux cellules en culture aide à préserver leur intégrité et peut également aider à conserver des fonctions plus différenciées pendant de plus longues périodes. D'autres études fondamentales ont permis d'approfondir la connaissance des besoins nutritionnels et hormonaux des cellules en culture, permettant de formuler de nouveaux milieux. Des progrès récents ont également été réalisés dans l'identification de matrices extracellulaires naturelles et artificielles sur lesquelles des cellules peuvent être cultivées. La culture de cellules sur ces différentes matrices peut avoir des effets profonds à la fois sur leur structure et leur fonction. Un avantage majeur dérivé de cette connaissance est la capacité de contrôler de manière complexe l'environnement des cellules en culture et d'examiner individuellement les effets de ces facteurs sur les processus cellulaires de base et sur leurs réponses à différents agents chimiques. En bref, ces systèmes peuvent fournir un excellent aperçu des mécanismes de toxicité spécifiques aux organes.
De nombreuses études de toxicité pour les organes cibles sont menées dans des cellules primaires, qui par définition sont fraîchement isolées d'un organe et présentent généralement une durée de vie finie en culture. Il y a de nombreux avantages à avoir des cultures primaires d'un seul type de cellule à partir d'un organe pour l'évaluation de la toxicité. D'un point de vue mécaniste, de telles cultures sont utiles pour étudier des cibles cellulaires spécifiques d'un produit chimique. Dans certains cas, deux ou plusieurs types de cellules d'un organe peuvent être cultivés ensemble, ce qui offre un avantage supplémentaire de pouvoir examiner les interactions cellule-cellule en réponse à une toxine. Certains systèmes de co-culture pour la peau ont été conçus de sorte qu'ils forment une structure tridimensionnelle ressemblant à la peau in vivo. Il est également possible de co-culturer des cellules de différents organes, par exemple le foie et les reins. Ce type de culture serait utile pour évaluer les effets propres aux cellules rénales d'une substance chimique qui doit être bioactivée dans le foie.
Les outils de biologie moléculaire ont également joué un rôle important dans le développement de lignées cellulaires continues qui peuvent être utiles pour les tests de toxicité sur les organes cibles. Ces lignées cellulaires sont générées en transfectant de l'ADN dans des cellules primaires. Dans la procédure de transfection, les cellules et l'ADN sont traités de sorte que l'ADN puisse être absorbé par les cellules. L'ADN provient généralement d'un virus et contient un gène ou des gènes qui, lorsqu'ils sont exprimés, permettent aux cellules de s'immortaliser (c'est-à-dire capables de vivre et de croître pendant de longues périodes de temps en culture). L'ADN peut également être modifié de manière à ce que le gène immortalisant soit contrôlé par un promoteur inductible. L'avantage de ce type de construction est que les cellules ne se diviseront que lorsqu'elles recevront le stimulus chimique approprié pour permettre l'expression du gène immortalisant. Un exemple d'une telle construction est le grand gène de l'antigène T du virus simien 40 (SV40) (le gène immortalisant), précédé de la région promotrice du gène de la métallothionéine, qui est induite par la présence d'un métal dans le milieu de culture. Ainsi, après que le gène est transfecté dans les cellules, les cellules peuvent être traitées avec de faibles concentrations de zinc pour stimuler le promoteur MT et activer l'expression du gène de l'antigène T. Dans ces conditions, les cellules prolifèrent. Lorsque le zinc est éliminé du milieu, les cellules arrêtent de se diviser et, dans des conditions idéales, reviennent à un état où elles expriment leurs fonctions spécifiques aux tissus.
La capacité de générer des cellules immortalisées combinée aux progrès de la technologie de la culture cellulaire a grandement contribué à la création de lignées cellulaires à partir de nombreux organes différents, notamment le cerveau, les reins et le foie. Cependant, avant que ces lignées cellulaires puissent être utilisées comme substitut des types de cellules authentiques, elles doivent être soigneusement caractérisées pour déterminer à quel point elles sont vraiment « normales ».
D'autres systèmes in vitro pour étudier la toxicité d'un organe cible impliquent une complexité croissante. Au fur et à mesure que les systèmes in vitro progressent en complexité, de la cellule unique à la culture d'organes entiers, ils deviennent plus comparables au milieu in vivo, mais en même temps, ils deviennent beaucoup plus difficiles à contrôler compte tenu du nombre accru de variables. Par conséquent, ce qui peut être gagné en passant à un niveau supérieur d'organisation peut être perdu dans l'incapacité du chercheur à contrôler l'environnement expérimental. Le tableau 1 compare certaines des caractéristiques de divers systèmes in vitro qui ont été utilisés pour étudier l'hépatotoxicité.
Tableau 1. Comparaison des systèmes in vitro pour les études d'hépatotoxicité
| Système | Complexité (niveau d'interaction) |
Capacité à conserver les fonctions spécifiques du foie | Durée potentielle de culture | Capacité à contrôler l'environnement |
| Lignées cellulaires immortalisées | certains de cellule à cellule (varie selon la lignée cellulaire) | médiocre à bon (varie selon la lignée cellulaire) | indéfini | excellent |
| Cultures primaires d'hépatocytes | cellule à cellule | passable à excellent (varie selon les conditions de culture) | jours à semaines | excellent |
| Co-cultures de cellules hépatiques | cellule à cellule (entre le même type de cellule et des types de cellules différents) | bon à excellent | semaines | excellent |
| Tranches de foie | cellule à cellule (parmi tous les types de cellules) | bon à excellent | heures en jours | Bien |
| Foie isolé et perfusé | cellule à cellule (parmi tous les types de cellules) et intra-organe | excellent | heures | juste |
Les tranches de tissu coupées avec précision sont utilisées plus largement pour les études toxicologiques. Il existe de nouveaux instruments disponibles qui permettent au chercheur de couper des tranches de tissu uniformes dans un environnement stérile. Les tranches de tissu offrent un certain avantage par rapport aux systèmes de culture cellulaire en ce que tous les types de cellules de l'organe sont présents et qu'ils conservent leur architecture in vivo et leur communication intercellulaire. Ainsi, des études in vitro peuvent être menées pour déterminer le type de cellule cible dans un organe ainsi que pour étudier la toxicité spécifique d'un organe cible. Un inconvénient des tranches est qu'elles dégénèrent rapidement après les premières 24 heures de culture, principalement en raison d'une mauvaise diffusion de l'oxygène vers les cellules à l'intérieur des tranches. Cependant, des études récentes ont indiqué qu'une aération plus efficace peut être obtenue par une rotation douce. Ceci, associé à l'utilisation d'un milieu plus complexe, permet aux tranches de survivre jusqu'à 96 heures.
Les explants de tissus sont similaires dans leur concept aux tranches de tissus et peuvent également être utilisés pour déterminer la toxicité de produits chimiques dans des organes cibles spécifiques. Les explants de tissus sont établis en prélevant un petit morceau de tissu (pour les études de tératogénicité, un embryon intact) et en le plaçant en culture pour une étude plus approfondie. Les cultures d'explants ont été utiles pour les études de toxicité à court terme, y compris l'irritation et la corrosivité de la peau, les études sur l'amiante dans la trachée et les études de neurotoxicité dans les tissus cérébraux.
Des organes perfusés isolés peuvent également être utilisés pour évaluer la toxicité des organes cibles. Ces systèmes offrent un avantage similaire à celui des tranches de tissu et des explants en ce que tous les types de cellules sont présents, mais sans le stress sur le tissu introduit par les manipulations impliquées dans la préparation des tranches. De plus, ils permettent le maintien des interactions intra-organes. Un inconvénient majeur est leur viabilité à court terme, ce qui limite leur utilisation pour les tests de toxicité in vitro. En termes de service d'alternative, ces cultures peuvent être considérées comme un raffinement puisque les animaux ne subissent pas les conséquences néfastes d'un traitement in vivo avec des substances toxiques. Cependant, leur utilisation ne diminue pas de manière significative le nombre d'animaux nécessaires.
En résumé, il existe plusieurs types de systèmes in vitro disponibles pour évaluer la toxicité des organes cibles. Il est possible d'acquérir de nombreuses informations sur les mécanismes de toxicité en utilisant une ou plusieurs de ces techniques. La difficulté reste de savoir extrapoler d'un système in vitro, qui représente une part relativement faible du processus toxicologique, à l'ensemble du processus se déroulant in vivo.
Tests in vitro pour l'irritation oculaire
Le test de toxicité sur l'animal entier le plus controversé du point de vue du bien-être animal est peut-être le test de Draize pour l'irritation des yeux, qui est effectué sur des lapins. Dans ce test, une petite dose fixe d'un produit chimique est placée dans l'un des yeux du lapin tandis que l'autre œil est utilisé comme témoin. Le degré d'irritation et d'inflammation est évalué à différents moments après l'exposition. Un effort important est fait pour développer des méthodologies pour remplacer ce test, qui a été critiqué non seulement pour des raisons humanitaires, mais aussi en raison de la subjectivité des observations et de la variabilité des résultats. Il est intéressant de noter que malgré les sévères critiques que le test de Draize a reçues, il s'est avéré remarquablement efficace pour prédire les irritants oculaires humains, en particulier les substances légèrement à modérément irritantes, qui sont difficiles à identifier par d'autres méthodes. Ainsi, les demandes sur les alternatives in vitro sont grandes.
La recherche d'alternatives au test de Draize est compliquée, même si elle devrait être couronnée de succès. De nombreuses alternatives in vitro et autres ont été développées et, dans certains cas, elles ont été mises en œuvre. Les alternatives de raffinement au test de Draize, qui, par définition, sont moins douloureuses ou stressantes pour les animaux, comprennent le test oculaire à faible volume, dans lequel de plus petites quantités de matériel de test sont placées dans les yeux des lapins, non seulement pour des raisons humanitaires, mais pour imiter plus fidèlement les quantités auxquelles les personnes peuvent être accidentellement exposées. Un autre raffinement est que les substances qui ont un pH inférieur à 2 ou supérieur à 11.5 ne sont plus testées sur les animaux car elles sont connues pour être sévèrement irritantes pour les yeux.
Entre 1980 et 1989, il y a eu une baisse estimée à 87 % du nombre de lapins utilisés pour les tests d'irritation oculaire des cosmétiques. Des tests in vitro ont été incorporés dans le cadre d'une approche de test à plusieurs niveaux pour provoquer cette vaste réduction des tests sur des animaux entiers. Cette approche est un processus en plusieurs étapes qui commence par un examen approfondi des données historiques sur l'irritation oculaire et une analyse physique et chimique du produit chimique à évaluer. Si ces deux processus ne fournissent pas suffisamment d'informations, une batterie de tests in vitro est réalisée. Les données supplémentaires obtenues à partir des tests in vitro pourraient alors être suffisantes pour évaluer la sécurité de la substance. Si ce n'est pas le cas, la dernière étape consisterait à effectuer des tests in vivo limités. Il est facile de voir comment cette approche peut éliminer ou au moins réduire considérablement le nombre d'animaux nécessaires pour prédire l'innocuité d'une substance d'essai.
La batterie de tests in vitro utilisée dans le cadre de cette stratégie de tests à plusieurs niveaux dépend des besoins de l'industrie en question. Les tests d'irritation oculaire sont effectués par une grande variété d'industries, des cosmétiques aux produits pharmaceutiques en passant par les produits chimiques industriels. Le type d'informations requises par chaque industrie varie et il n'est donc pas possible de définir une seule batterie de tests in vitro. Une batterie de tests est généralement conçue pour évaluer cinq paramètres : la cytotoxicité, les modifications de la physiologie et de la biochimie des tissus, les relations quantitatives structure-activité, les médiateurs de l'inflammation, ainsi que la récupération et la réparation. Un exemple de test de cytotoxicité, qui est une cause possible d'irritation, est le test au rouge neutre utilisant des cellules en culture (voir ci-dessus). Les modifications de la physiologie cellulaire et de la biochimie résultant de l'exposition à un produit chimique peuvent être dosées dans des cultures de cellules épithéliales cornéennes humaines. Alternativement, les enquêteurs ont également utilisé des globes oculaires de bovin ou de poulet intacts ou disséqués provenant d'abattoirs. De nombreux paramètres mesurés dans ces cultures d'organes entiers sont les mêmes que ceux mesurés in vivo, tels que l'opacité cornéenne et le gonflement cornéen.
L'inflammation est souvent une composante des lésions oculaires induites par des produits chimiques, et il existe un certain nombre de tests disponibles pour examiner ce paramètre. Divers dosages biochimiques détectent la présence de médiateurs libérés au cours du processus inflammatoire tels que l'acide arachidonique et les cytokines. La membrane chorioallantoïque (CAM) de l'œuf de poule peut également être utilisée comme indicateur d'inflammation. Dans le test CAM, un petit morceau de la coquille d'un embryon de poulet de dix à 14 jours est retiré pour exposer le CAM. Le produit chimique est ensuite appliqué sur la CAM et les signes d'inflammation, tels qu'une hémorragie vasculaire, sont notés à divers moments par la suite.
L'un des processus in vivo les plus difficiles à évaluer in vitro est la récupération et la réparation des lésions oculaires. Un instrument nouvellement développé, le microphysiomètre au silicium, mesure de petits changements dans le pH extracellulaire et peut être utilisé pour surveiller les cellules cultivées en temps réel. Cette analyse s'est avérée assez bien corrélée avec la récupération in vivo et a été utilisée comme test in vitro pour ce processus. Ceci a été un bref aperçu des types de tests utilisés comme alternatives au test de Draize pour l'irritation oculaire. Il est probable qu'au cours des prochaines années, une série complète de batteries de tests in vitro sera définie et chacune sera validée pour son objectif spécifique.
Validation
La clé de l'acceptation réglementaire et de la mise en œuvre des méthodologies de test in vitro est la validation, le processus par lequel la crédibilité d'un test candidat est établie dans un but spécifique. Des efforts pour définir et coordonner le processus de validation ont été faits tant aux États-Unis qu'en Europe. L'Union européenne a créé le Centre européen pour la validation des méthodes alternatives (ECVAM) en 1993 pour y coordonner les efforts et interagir avec des organisations américaines telles que le Johns Hopkins Center for Alternatives to Animal Testing (CAAT), un centre universitaire aux États-Unis. , et l'Interagency Coordinating Committee for the Validation of Alternative Methods (ICCVAM), composé de représentants des National Institutes of Health, de l'Environmental Protection Agency des États-Unis, de la Food and Drug Administration des États-Unis et de la Consumer Products Safety Commission.
La validation des tests in vitro nécessite une organisation et une planification importantes. Il doit y avoir un consensus entre les régulateurs gouvernementaux et les scientifiques industriels et universitaires sur les procédures acceptables, et une surveillance suffisante par un conseil consultatif scientifique pour s'assurer que les protocoles respectent les normes établies. Les études de validation doivent être réalisées dans une série de laboratoires de référence à l'aide d'ensembles étalonnés de produits chimiques provenant d'une banque de produits chimiques et de cellules ou de tissus provenant d'une source unique. La répétabilité intralaboratoire et la reproductibilité interlaboratoire d'un test candidat doivent être démontrées et les résultats soumis à une analyse statistique appropriée. Une fois les résultats des différentes composantes des études de validation compilés, le comité consultatif scientifique peut faire des recommandations sur la validité du ou des tests candidats dans un but précis. De plus, les résultats des études devraient être publiés dans des revues à comité de lecture et placés dans une base de données.
La définition du processus de validation est actuellement un travail en cours. Chaque nouvelle étude de validation apportera des informations utiles à la conception de l'étude suivante. La communication et la coopération internationales sont essentielles pour le développement rapide d'une série de protocoles largement acceptables, en particulier compte tenu de l'urgence accrue imposée par l'adoption de la directive CE sur les cosmétiques. Cette législation peut en effet donner l'impulsion nécessaire pour qu'un sérieux effort de validation soit entrepris. Ce n'est qu'après l'achèvement de ce processus que l'acceptation des méthodes in vitro par les diverses communautés réglementaires peut commencer.
Conclusion
Cet article a fourni un large aperçu de l'état actuel des tests de toxicité in vitro. La science de la toxicologie in vitro est relativement jeune, mais elle connaît une croissance exponentielle. Le défi pour les années à venir est d'intégrer les connaissances mécanistes générées par les études cellulaires et moléculaires dans le vaste inventaire des données in vivo pour fournir une description plus complète des mécanismes toxicologiques ainsi que pour établir un paradigme par lequel les données in vitro peuvent être utilisées. prédire la toxicité in vivo. Ce ne sera que grâce aux efforts concertés des toxicologues et des représentants gouvernementaux que la valeur intrinsèque de ces méthodes in vitro pourra être réalisée.
Évaluation de la toxicité génétique
L'évaluation de la toxicité génétique est l'évaluation des agents pour leur capacité à induire l'un des trois types généraux de changements (mutations) dans le matériel génétique (ADN) : gène, chromosomique et génomique. Dans des organismes tels que les humains, les gènes sont composés d'ADN, qui se compose d'unités individuelles appelées bases nucléotidiques. Les gènes sont disposés dans des structures physiques discrètes appelées chromosomes. La génotoxicité peut entraîner des effets importants et irréversibles sur la santé humaine. Les dommages génotoxiques sont une étape critique dans l'induction du cancer et peuvent également être impliqués dans l'induction de malformations congénitales et de mort fœtale. Les trois classes de mutations mentionnées ci-dessus peuvent se produire dans l'un ou l'autre des deux types de tissus possédés par des organismes tels que les humains : les spermatozoïdes ou les ovules (cellules germinales) et le tissu restant (cellules somatiques).
Les tests qui mesurent la mutation génique sont ceux qui détectent la substitution, l'addition ou la suppression de nucléotides dans un gène. Les tests qui mesurent la mutation chromosomique sont ceux qui détectent les cassures ou les réarrangements chromosomiques impliquant un ou plusieurs chromosomes. Les tests qui mesurent la mutation génomique sont ceux qui détectent les changements dans le nombre de chromosomes, une condition appelée aneuploïdie. L'évaluation de la toxicité génétique a considérablement évolué depuis la mise au point par Herman Muller en 1927 du premier test de détection d'agents génotoxiques (mutagènes). Depuis lors, plus de 200 tests ont été développés pour mesurer les mutations de l'ADN ; cependant, moins de dix tests sont couramment utilisés aujourd'hui pour l'évaluation de la toxicité génétique. Cet article passe en revue ces essais, décrit ce qu'ils mesurent et explore le rôle de ces essais dans l'évaluation de la toxicité.
Identification des risques de cancerAvant le développement du Domaine de la toxicologie génétique
La toxicologie génétique est devenue une partie intégrante du processus global d'évaluation des risques et s'est récemment imposée comme un prédicteur fiable de l'activité cancérigène. Cependant, avant le développement de la toxicologie génétique (avant 1970), d'autres méthodes étaient et sont toujours utilisées pour identifier les risques potentiels de cancer chez l'homme. Il existe six grandes catégories de méthodes actuellement utilisées pour identifier les risques de cancer chez l'homme : les études épidémiologiques, les bioessais in vivo à long terme, les bioessais in vivo à moyen terme, les bioessais in vivo et in vitro à court terme, l'intelligence artificielle (structure-activité), et l'inférence basée sur le mécanisme.
Le tableau 1 donne les avantages et les inconvénients de ces méthodes.
Tableau 1. Avantages et inconvénients des méthodes actuelles d'identification des risques de cancer chez l'homme
| Avantages | Inconvénients | |
| Les études épidémiologiques | (1) les humains sont les indicateurs ultimes de la maladie ; (2) évaluer les populations sensibles ou sensibles ; (3) cohortes d'exposition professionnelle; (4) alertes sentinelles environnementales |
(1) généralement rétrospectif (certificats de décès, biais de rappel, etc.) ; (2) insensible, coûteux, long ; (3) des données d'exposition fiables parfois indisponibles ou difficiles à obtenir ; (4) expositions combinées, multiples et complexes; manque de cohortes de contrôle appropriées; (5) les expériences sur les humains ne sont pas faites ; (6) détection du cancer, pas prévention |
| Essais biologiques in vivo à long terme | (1) évaluations prospectives et rétrospectives (validation) ; (2) excellente corrélation avec les carcinogènes humains identifiés; (3) niveaux et conditions d'exposition connus; (4) identifie la toxicité chimique et les effets cancérigènes ; (5) des résultats obtenus assez rapidement ; (6) comparaisons qualitatives entre classes chimiques; (7) systèmes biologiques intégratifs et interactifs étroitement liés aux humains | (1) rarement reproduit, gourmand en ressources ; (3) des installations limitées adaptées à de telles expériences ; (4) débat sur l'extrapolation des espèces; (5) les expositions utilisées sont souvent à des niveaux bien supérieurs à ceux subis par les humains; (6) l'exposition à un seul produit chimique n'imite pas les expositions humaines, qui sont généralement à plusieurs produits chimiques simultanément |
| Essais biologiques in vivo et in vitro à moyen et court terme | (1) plus rapide et moins cher que les autres tests ; (2) de grands échantillons facilement reproductibles ; (3) les points limites biologiquement significatifs sont mesurés (mutation, etc.); (4) peuvent être utilisés comme essais de dépistage pour sélectionner des produits chimiques pour des essais biologiques à long terme |
(1) in vitro pas entièrement prédictif d'in vivo; (2) généralement spécifiques à un organisme ou à un organe ; (3) puissances non comparables à des animaux entiers ou à des humains |
| Associations structure chimique–activité biologique | (1) relativement facile, rapide et peu coûteux ; (2) fiable pour certaines classes chimiques (par exemple, les colorants nitrosamines et benzidine); (3) développé à partir de données biologiques mais non dépendant d'expérimentations biologiques supplémentaires | (1) non « biologique » ; (2) de nombreuses exceptions aux règles formulées; (3) rétrospective et rarement (mais devenant) prospective |
| Inférences basées sur le mécanisme | (1) raisonnablement précis pour certaines classes de produits chimiques ; (2) permet d'affiner les hypothèses ; (3) peut orienter les évaluations des risques vers les populations sensibles | (1) mécanismes de carcinogenèse chimique indéfinis, multiples et probablement chimiques ou spécifiques à une classe ; (2) peut ne pas mettre en évidence les exceptions aux mécanismes généraux |
Justification et fondement conceptuel des tests de toxicologie génétique
Bien que les types et le nombre exacts de tests utilisés pour l'évaluation de la toxicité génétique évoluent constamment et varient d'un pays à l'autre, les plus courants incluent des tests pour (1) la mutation génique dans les bactéries et/ou les cellules de mammifères cultivées et (2) la mutation chromosomique dans des cellules de mammifères cultivées et/ou de la moelle osseuse chez des souris vivantes. Certains des tests de cette deuxième catégorie peuvent également détecter l'aneuploïdie. Bien que ces tests ne détectent pas les mutations dans les cellules germinales, ils sont principalement utilisés en raison du coût supplémentaire et de la complexité de la réalisation des tests sur les cellules germinales. Néanmoins, les tests sur les cellules germinales chez la souris sont utilisés lorsque des informations sur les effets sur les cellules germinales sont souhaitées.
Des études systématiques sur une période de 25 ans (1970-1995), en particulier au US National Toxicology Program en Caroline du Nord, ont abouti à l'utilisation d'un nombre discret de tests pour détecter l'activité mutagène des agents. La raison d'être de l'évaluation de l'utilité des tests reposait sur leur capacité à détecter des agents qui causent le cancer chez les rongeurs et qui sont soupçonnés de causer le cancer chez l'homme (c.-à-d., des agents cancérigènes). En effet, des études menées au cours des dernières décennies ont indiqué que les cellules cancéreuses contiennent des mutations dans certains gènes et que de nombreux agents cancérigènes sont également mutagènes. Ainsi, les cellules cancéreuses sont considérées comme contenant des mutations des cellules somatiques et la carcinogenèse est considérée comme un type de mutagenèse des cellules somatiques.
Les tests de toxicité génétique les plus couramment utilisés aujourd'hui ont été sélectionnés non seulement en raison de leur grande base de données, de leur coût relativement faible et de leur facilité d'exécution, mais aussi parce qu'il a été démontré qu'ils détectent de nombreux cancérogènes chez les rongeurs et, par présomption, chez l'homme. Par conséquent, des tests de toxicité génétique sont utilisés pour prédire la cancérogénicité potentielle des agents.
Un développement conceptuel et pratique important dans le domaine de la toxicologie génétique a été la reconnaissance que de nombreux cancérogènes étaient modifiés par des enzymes dans le corps, créant des formes altérées (métabolites) qui étaient souvent la forme cancérigène et mutagène ultime de la substance chimique mère. Pour dupliquer ce métabolisme dans une boîte de Pétri, Heinrich Malling a montré que l'inclusion d'une préparation de foie de rongeur contenait de nombreuses enzymes nécessaires pour effectuer cette conversion ou activation métabolique. Ainsi, de nombreux tests de toxicité génétique effectués dans des boîtes ou des tubes (in vitro) emploient l'addition de préparations enzymatiques similaires. Les préparations simples sont appelées mélange S9 et les préparations purifiées sont appelées microsomes. Certaines cellules bactériennes et mammifères ont maintenant été génétiquement modifiées pour contenir certains des gènes de rongeurs ou d'humains qui produisent ces enzymes, réduisant ainsi la nécessité d'ajouter un mélange S9 ou des microsomes.
Dosages et techniques de toxicologie génétique
Les principaux systèmes bactériens utilisés pour le dépistage de la toxicité génétique sont le test de mutagénicité de Salmonella (Ames) et, dans une bien moindre mesure, la souche WP2 de Escherichia coli. Des études menées au milieu des années 1980 ont indiqué que l'utilisation de seulement deux souches du système Salmonella (TA98 et TA100) était suffisante pour détecter environ 90 % des mutagènes connus de Salmonella. Ainsi, ces deux souches sont utilisées pour la plupart des objectifs de dépistage ; cependant, diverses autres souches sont disponibles pour des tests plus approfondis.
Ces dosages sont effectués de diverses manières, mais deux procédures générales sont les dosages d'incorporation sur plaque et de suspension liquide. Dans le test d'incorporation sur plaque, les cellules, le produit chimique d'essai et (le cas échéant) le S9 sont ajoutés ensemble dans une gélose liquéfiée et versés sur la surface d'une boîte de Pétri d'agar. La gélose supérieure durcit en quelques minutes et les plaques sont incubées pendant deux à trois jours, après quoi les cellules mutantes se sont développées pour former des amas de cellules détectables visuellement appelées colonies, qui sont ensuite comptées. Le milieu gélosé contient des agents sélectifs ou est composé d'ingrédients tels que seules les cellules nouvellement mutées se développeront. Le test d'incubation liquide est similaire, sauf que les cellules, l'agent de test et S9 sont incubés ensemble dans un liquide qui ne contient pas d'agar liquéfié, puis les cellules sont lavées sans l'agent de test et S9 et ensemencées sur l'agar.
Les mutations dans les cellules de mammifères en culture sont principalement détectées dans l'un des deux gènes suivants : hprt ainsi que tk. Comme pour les tests bactériens, les lignées cellulaires de mammifères (développées à partir de cellules de rongeurs ou humaines) sont exposées à l'agent de test dans des boîtes ou des tubes de culture en plastique, puis sont ensemencées dans des boîtes de culture contenant un milieu avec un agent sélectif qui permet uniquement aux cellules mutantes de se développer. . Les tests utilisés à cette fin comprennent le CHO/HPRT, le TK6 et le lymphome de souris L5178Y/TK+/- dosages. D'autres lignées cellulaires contenant diverses mutations de réparation de l'ADN ainsi que certains gènes humains impliqués dans le métabolisme sont également utilisées. Ces systèmes permettent la récupération de mutations au sein du gène (mutation génique) ainsi que de mutations impliquant des régions du chromosome flanquant le gène (mutation chromosomique). Cependant, ce dernier type de mutation est beaucoup plus récupéré par la tk systèmes de gènes que par le hprt systèmes de gènes en raison de l'emplacement du tk .
Semblable au test d'incubation liquide pour la mutagénicité bactérienne, les tests de mutagénicité sur les cellules de mammifères impliquent généralement l'exposition des cellules dans des boîtes ou des tubes de culture en présence de l'agent de test et de S9 pendant plusieurs heures. Les cellules sont ensuite lavées, cultivées pendant plusieurs jours supplémentaires pour permettre la dégradation des produits géniques normaux (de type sauvage) et l'expression et l'accumulation des produits géniques nouvellement mutants, puis elles sont ensemencées dans un milieu contenant un agent sélectif qui permet seules les cellules mutantes se développent. Comme les tests bactériens, les cellules mutantes se développent en colonies visuellement détectables qui sont ensuite comptées.
La mutation chromosomique est identifiée principalement par des tests cytogénétiques, qui impliquent d'exposer des rongeurs et/ou des cellules de rongeurs ou humaines dans des boîtes de culture à un produit chimique d'essai, permettant à une ou plusieurs divisions cellulaires de se produire, de colorer les chromosomes, puis d'examiner visuellement les chromosomes au microscope. détecter des altérations de la structure ou du nombre de chromosomes. Bien qu'une variété de paramètres puissent être examinés, les deux qui sont actuellement acceptés par les organismes de réglementation comme étant les plus significatifs sont les aberrations chromosomiques et une sous-catégorie appelée micronoyaux.
Une formation et une expertise considérables sont nécessaires pour évaluer la présence d'aberrations chromosomiques dans les cellules, ce qui en fait une procédure coûteuse en temps et en argent. En revanche, les micronoyaux nécessitent peu de formation et leur détection peut être automatisée. Les micronoyaux apparaissent sous forme de petits points dans la cellule qui sont distincts du noyau, qui contient les chromosomes. Les micronoyaux résultent soit d'une rupture chromosomique, soit d'une aneuploïdie. En raison de la facilité de notation des micronoyaux par rapport aux aberrations chromosomiques, et parce que des études récentes indiquent que les agents qui induisent des aberrations chromosomiques dans la moelle osseuse des souris vivantes induisent généralement des micronoyaux dans ce tissu, les micronoyaux sont maintenant couramment mesurés comme une indication de la capacité d'un agent pour induire la mutation chromosomique.
Bien que les tests sur les cellules germinales soient utilisés beaucoup moins fréquemment que les autres tests décrits ci-dessus, ils sont indispensables pour déterminer si un agent présente un risque pour les cellules germinales, dont les mutations peuvent entraîner des effets sur la santé des générations suivantes. Les tests de cellules germinales les plus couramment utilisés sont chez la souris et impliquent des systèmes qui détectent (1) les translocations héréditaires (échanges) entre les chromosomes (test de translocation héréditaire), (2) les mutations génétiques ou chromosomiques impliquant des gènes spécifiques (locus visible ou biochimique spécifique). tests) et (3) les mutations qui affectent la viabilité (dosage létal dominant). Comme pour les tests sur les cellules somatiques, l'hypothèse de travail avec les tests sur les cellules germinales est que les agents positifs dans ces tests sont présumés être des mutagènes potentiels des cellules germinales humaines.
Situation actuelle et perspectives d'avenir
Des études récentes ont indiqué que seulement trois éléments d'information étaient nécessaires pour détecter environ 90 % d'un ensemble de 41 cancérogènes chez les rongeurs (c.-à-d., cancérogènes humains présumés et mutagènes des cellules somatiques). Celles-ci comprenaient (1) la connaissance de la structure chimique de l'agent, en particulier s'il contient des fractions électrophiles (voir la section sur les relations structure-activité) ; (2) Données sur la mutagénicité de Salmonella ; et (3) les données d'un test de toxicité chronique de 90 jours chez les rongeurs (souris et rats). En effet, pratiquement tous les cancérogènes humains déclarés par le CIRC sont détectables en tant que mutagènes en utilisant uniquement le test Salmonella et le test du micronoyau de moelle osseuse de souris. L'utilisation de ces essais de mutagénicité pour détecter des cancérogènes humains potentiels est étayée par la découverte que la plupart des cancérogènes pour l'homme sont cancérigènes à la fois pour les rats et les souris (cancérigènes trans-espèces) et que la plupart des cancérogènes trans-espèces sont mutagènes pour Salmonella et/ou induisent des micronoyaux. dans la moelle osseuse de souris.
Avec les progrès de la technologie de l'ADN, le projet du génome humain et une meilleure compréhension du rôle de la mutation dans le cancer, de nouveaux tests de génotoxicité sont en cours de développement et seront probablement intégrés aux procédures de dépistage standard. Parmi ceux-ci figurent l'utilisation de cellules transgéniques et de rongeurs. Les systèmes transgéniques sont ceux dans lesquels un gène d'une autre espèce a été introduit dans une cellule ou un organisme. Par exemple, des souris transgéniques sont maintenant utilisées à titre expérimental pour permettre la détection d'une mutation dans n'importe quel organe ou tissu de l'animal, sur la base de l'introduction d'un gène bactérien dans la souris. Des cellules bactériennes, telles que Salmonella, et des cellules de mammifères (y compris des lignées cellulaires humaines) sont désormais disponibles et contiennent des gènes impliqués dans le métabolisme d'agents cancérigènes/mutagènes, tels que les gènes P450. L'analyse moléculaire des mutations réelles induites dans le trans-gène chez les rongeurs transgéniques, ou dans les gènes natifs tels que hprt, ou les gènes cibles au sein de Salmonella peuvent maintenant être analysés, de sorte que la nature exacte des mutations induites par les produits chimiques puisse être déterminée, fournissant des informations sur le mécanisme d'action du produit chimique et permettant des comparaisons avec des mutations chez des humains présumés exposés à l'agent .
Les progrès moléculaires de la cytogénétique permettent maintenant une évaluation plus détaillée des mutations chromosomiques. Celles-ci incluent l'utilisation de sondes (petits morceaux d'ADN) qui se fixent (s'hybrident) à des gènes spécifiques. Des réarrangements de gènes sur le chromosome peuvent alors être révélés par la localisation altérée des sondes, qui sont fluorescentes et facilement visualisables sous forme de secteurs colorés sur les chromosomes. Le test d'électrophorèse sur gel unicellulaire pour la rupture de l'ADN (communément appelé le test «comète») permet la détection des ruptures d'ADN dans des cellules individuelles et peut devenir un outil extrêmement utile en combinaison avec des techniques cytogénétiques pour détecter les dommages chromosomiques.
Après de nombreuses années d'utilisation et la génération d'une base de données importante et systématiquement développée, l'évaluation de la toxicité génétique peut désormais être effectuée avec seulement quelques tests pour un coût relativement faible dans un court laps de temps (quelques semaines). Les données produites peuvent être utilisées pour prédire la capacité d'un agent à être un rongeur et, par présomption, un cancérigène humain/mutagène des cellules somatiques. Une telle capacité permet de limiter l'introduction dans l'environnement d'agents mutagènes et cancérigènes et de développer des agents alternatifs non mutagènes. Les études futures devraient conduire à des méthodes encore meilleures avec une plus grande prédictivité que les tests actuels.
Biomarqueurs
Le mot biomarqueur est l'abréviation de marqueur biologique, un terme qui fait référence à un événement mesurable se produisant dans un système biologique, tel que le corps humain. Cet événement est alors interprété comme le reflet, ou le marqueur, d'un état plus général de l'organisme ou de l'espérance de vie. En santé au travail, un biomarqueur est généralement utilisé comme indicateur de l'état de santé ou du risque de maladie.
Les biomarqueurs sont utilisés pour des études in vitro et in vivo qui peuvent inclure des humains. Habituellement, trois types spécifiques de marqueurs biologiques sont identifiés. Bien que quelques biomarqueurs puissent être difficiles à classer, ils sont généralement séparés en biomarqueurs d'exposition, biomarqueurs d'effet ou biomarqueurs de sensibilité (voir tableau 1).
Tableau 1. Exemples de biomarqueurs d'exposition ou biomarqueurs d'effet utilisés dans les études toxicologiques en santé au travail
| Échantillon | Mesure | Objectif |
| Biomarqueurs d'exposition | ||
| Tissu adipeux | dioxine | Exposition aux dioxines |
| sanguins | Plomb | Exposition au plomb |
| Greffe Osseuse | Aluminium | Exposition à l'aluminium |
| Souffle expiré | Toluène | Exposition au toluène |
| Implants | Mercury | Exposition au méthylmercure |
| Sérum | Benzène | Exposition au benzène |
| Urine | Phénol | Exposition au benzène |
| Biomarqueurs d'effet | ||
| sanguins | Carboxyhémoglobine | Exposition au monoxyde de carbone |
| des globules rouges | Zinc-protoporphyrine | Exposition au plomb |
| Sérum | Cholinestérase | Exposition aux organophosphorés |
| Urine | Microglobulines | Exposition néphrotoxique |
| Les globules blancs | Adduits à l'ADN | Exposition mutagène |
Avec un degré de validité acceptable, les biomarqueurs peuvent être utilisés à plusieurs fins. Sur une base individuelle, un biomarqueur peut être utilisé pour étayer ou réfuter un diagnostic d'un type particulier d'empoisonnement ou d'un autre effet indésirable d'origine chimique. Chez un sujet sain, un biomarqueur peut également refléter l'hypersensibilité individuelle à des expositions chimiques spécifiques et peut donc servir de base pour la prédiction des risques et le conseil. Dans les groupes de travailleurs exposés, certains biomarqueurs d'exposition peuvent être appliqués pour évaluer le degré de conformité aux réglementations antipollution ou l'efficacité des efforts de prévention en général.
Biomarqueurs d'exposition
Un biomarqueur d'exposition peut être un composé exogène (ou un métabolite) dans le corps, un produit interactif entre le composé (ou le métabolite) et un composant endogène, ou un autre événement lié à l'exposition. Le plus souvent, les biomarqueurs des expositions à des composés stables, tels que les métaux, comprennent des mesures des concentrations de métaux dans des échantillons appropriés, tels que le sang, le sérum ou l'urine. Avec les produits chimiques volatils, leur concentration dans l'air expiré (après inhalation d'air non contaminé) peut être évaluée. Si le composé est métabolisé dans l'organisme, un ou plusieurs métabolites peuvent être choisis comme biomarqueur de l'exposition ; les métabolites sont souvent déterminés dans des échantillons d'urine.
Les méthodes modernes d'analyse peuvent permettre la séparation d'isomères ou de congénères de composés organiques et la détermination de la spéciation de composés métalliques ou des rapports isotopiques de certains éléments. Des analyses sophistiquées permettent de déterminer les changements dans la structure de l'ADN ou d'autres macromolécules provoqués par la liaison avec des produits chimiques réactifs. Ces techniques avancées gagneront sans aucun doute considérablement en importance pour les applications dans les études de biomarqueurs, et des limites de détection plus basses et une meilleure validité analytique rendront probablement ces biomarqueurs encore plus utiles.
Des développements particulièrement prometteurs ont eu lieu avec des biomarqueurs d'exposition à des produits chimiques mutagènes. Ces composés sont réactifs et peuvent former des adduits avec des macromolécules, telles que des protéines ou de l'ADN. Des adduits d'ADN peuvent être détectés dans les globules blancs ou des biopsies tissulaires, et des fragments d'ADN spécifiques peuvent être excrétés dans l'urine. Par exemple, l'exposition à l'oxyde d'éthylène entraîne des réactions avec les bases de l'ADN et, après excision de la base endommagée, la N-7-(2-hydroxyéthyl)guanine sera éliminée dans les urines. Certains adduits peuvent ne pas se référer directement à une exposition particulière. Par exemple, la 8-hydroxy-2´-désoxyguanosine reflète les dommages oxydatifs de l'ADN, et cette réaction peut être déclenchée par plusieurs composés chimiques, dont la plupart induisent également une peroxydation lipidique.
D'autres macromolécules peuvent également être modifiées par formation d'adduits ou oxydation. D'un intérêt particulier, de tels composés réactifs peuvent générer des adduits d'hémoglobine qui peuvent être déterminés en tant que biomarqueurs d'exposition aux composés. L'avantage est que de grandes quantités d'hémoglobine peuvent être obtenues à partir d'un échantillon de sang et, étant donné la durée de vie de quatre mois des globules rouges, les adduits formés avec les acides aminés de la protéine indiqueront l'exposition totale pendant cette période.
Les adduits peuvent être déterminés par des techniques sensibles telles que la chromatographie lipidique à haute performance, et certaines méthodes immunologiques sont également disponibles. En général, les méthodes analytiques sont nouvelles, coûteuses et nécessitent un développement et une validation supplémentaires. Une meilleure sensibilité peut être obtenue en utilisant le 32Test de post-marquage P, qui est une indication non spécifique que des dommages à l'ADN ont eu lieu. Toutes ces techniques sont potentiellement utiles pour la surveillance biologique et ont été appliquées dans un nombre croissant d'études. Cependant, des méthodes analytiques plus simples et plus sensibles sont nécessaires. Compte tenu de la spécificité limitée de certaines méthodes à de faibles niveaux d'exposition, le tabagisme ou d'autres facteurs peuvent avoir un impact significatif sur les résultats de mesure, entraînant ainsi des difficultés d'interprétation.
L'exposition à des composés mutagènes ou à des composés qui sont métabolisés en mutagènes peut également être déterminée en évaluant la mutagénicité de l'urine d'un individu exposé. L'échantillon d'urine est incubé avec une souche de bactéries dans laquelle une mutation ponctuelle spécifique est exprimée d'une manière qui peut être facilement mesurée. Si des produits chimiques mutagènes sont présents dans l'échantillon d'urine, un taux accru de mutations se produira dans les bactéries.
Les biomarqueurs d'exposition doivent être évalués au regard de la variation temporelle de l'exposition et de la relation aux différents compartiments. Ainsi, la ou les périodes de temps représentées par le biomarqueur, c'est-à-dire la mesure dans laquelle la mesure du biomarqueur reflète l'exposition ou les expositions passées et/ou la charge corporelle accumulée, doivent être déterminées à partir des données toxicocinétiques afin d'interpréter le résultat. En particulier, le degré auquel le biomarqueur indique une rétention dans des organes cibles spécifiques doit être pris en compte. Bien que les échantillons de sang soient souvent utilisés pour les études de biomarqueurs, le sang périphérique n'est généralement pas considéré comme un compartiment en tant que tel, bien qu'il agisse comme un milieu de transport entre les compartiments. La mesure dans laquelle la concentration dans le sang reflète les niveaux dans différents organes varie considérablement entre les différents produits chimiques et dépend généralement aussi de la durée de l'exposition ainsi que du temps écoulé depuis l'exposition.
Parfois, ce type de preuve est utilisé pour classer un biomarqueur comme un indicateur de dose absorbée (totale) ou un indicateur de dose efficace (c'est-à-dire la quantité qui a atteint le tissu cible). Par exemple, l'exposition à un solvant particulier peut être évaluée à partir de données sur la concentration réelle du solvant dans le sang à un moment particulier après l'exposition. Cette mesure reflétera la quantité de solvant qui a été absorbée par le corps. Une partie de la quantité absorbée sera expirée en raison de la pression de vapeur du solvant. En circulant dans le sang, le solvant interagira avec divers composants du corps et finira par être dégradé par les enzymes. Le résultat des processus métaboliques peut être évalué en déterminant des acides mercapturiques spécifiques produits par conjugaison avec le glutathion. L'excrétion cumulée des acides mercapturiques peut mieux refléter la dose efficace que la concentration sanguine.
Les événements de la vie, tels que la reproduction et la sénescence, peuvent affecter la distribution d'un produit chimique. La distribution des produits chimiques dans le corps est considérablement affectée par la grossesse, et de nombreux produits chimiques peuvent traverser la barrière placentaire, provoquant ainsi une exposition du fœtus. La lactation peut entraîner l'excrétion de produits chimiques liposolubles, entraînant ainsi une diminution de la rétention chez la mère ainsi qu'une augmentation de l'absorption par le nourrisson. Pendant la perte de poids ou le développement de l'ostéoporose, des produits chimiques stockés peuvent être libérés, ce qui peut alors entraîner une exposition «endogène» renouvelée et prolongée des organes cibles. D'autres facteurs peuvent affecter l'absorption individuelle, le métabolisme, la rétention et la distribution des composés chimiques, et certains biomarqueurs de sensibilité sont disponibles (voir ci-dessous).
Biomarqueurs d'effet
Un marqueur d'effet peut être un composant endogène, ou une mesure de la capacité fonctionnelle, ou un autre indicateur de l'état ou de l'équilibre du corps ou du système organique, tel qu'affecté par l'exposition. De tels marqueurs d'effets sont généralement des indicateurs précliniques d'anomalies.
Ces biomarqueurs peuvent être spécifiques ou non spécifiques. Les biomarqueurs spécifiques sont utiles car ils indiquent un effet biologique d'une exposition particulière, fournissant ainsi des preuves qui peuvent potentiellement être utilisées à des fins préventives. Les biomarqueurs non spécifiques n'indiquent pas une cause individuelle de l'effet, mais ils peuvent refléter l'effet total intégré dû à une exposition mixte. Les deux types de biomarqueurs peuvent donc être d'une utilité considérable en santé au travail.
Il n'y a pas de distinction claire entre les biomarqueurs d'exposition et les biomarqueurs d'effet. Par exemple, on pourrait dire que la formation d'adduits reflète un effet plutôt que l'exposition. Cependant, les biomarqueurs d'effet indiquent généralement des changements dans les fonctions des cellules, des tissus ou de l'ensemble du corps. Certains chercheurs incluent des changements brutaux, tels qu'une augmentation du poids du foie des animaux de laboratoire exposés ou une diminution de la croissance chez les enfants, comme biomarqueurs d'effet. Aux fins de la santé au travail, les biomarqueurs d'effets devraient être limités à ceux qui indiquent des modifications biochimiques subcliniques ou réversibles, telles que l'inhibition des enzymes. Le biomarqueur d'effet le plus fréquemment utilisé est probablement l'inhibition de la cholinestérase causée par certains insecticides, c'est-à-dire les organophosphorés et les carbamates. Dans la plupart des cas, cet effet est entièrement réversible et l'inhibition enzymatique reflète l'exposition totale à ce groupe particulier d'insecticides.
Certaines expositions n'entraînent pas d'inhibition enzymatique mais plutôt une activité accrue d'une enzyme. C'est le cas de plusieurs enzymes appartenant à la famille P450 (voir « Déterminants génétiques de la réponse toxique »). Ils peuvent être induits par des expositions à certains solvants et hydrocarbures polyaromatiques (HAP). Étant donné que ces enzymes sont principalement exprimées dans les tissus à partir desquels une biopsie peut être difficile à obtenir, l'activité enzymatique est déterminée indirectement in vivo en administrant un composé qui est métabolisé par cette enzyme particulière, puis le produit de dégradation est mesuré dans l'urine ou le plasma.
D'autres expositions peuvent induire la synthèse d'une protéine protectrice dans l'organisme. Le meilleur exemple est probablement la métallothionéine, qui lie le cadmium et favorise l'excrétion de ce métal ; l'exposition au cadmium est l'un des facteurs qui entraînent une expression accrue du gène de la métallothionéine. Des protéines protectrices similaires peuvent exister mais n'ont pas encore été suffisamment explorées pour être acceptées comme biomarqueurs. Parmi les candidats à une utilisation possible en tant que biomarqueurs figurent les protéines dites de stress, appelées à l'origine protéines de choc thermique. Ces protéines sont générées par une gamme d'organismes différents en réponse à une variété d'expositions nocives.
Les dommages oxydatifs peuvent être évalués en déterminant la concentration de malondialdéhyde dans le sérum ou l'exhalation d'éthane. De même, l'excrétion urinaire de protéines de faible poids moléculaire, comme l'albumine, peut être utilisée comme biomarqueur d'une atteinte rénale précoce. Plusieurs paramètres couramment utilisés en pratique clinique (par exemple, les taux sériques d'hormones ou d'enzymes) peuvent également être utiles en tant que biomarqueurs. Cependant, bon nombre de ces paramètres peuvent ne pas être suffisamment sensibles pour détecter une déficience précoce.
Un autre groupe de paramètres d'effet concerne les effets génotoxiques (modifications de la structure des chromosomes). De tels effets peuvent être détectés par microscopie des globules blancs qui subissent une division cellulaire. De graves dommages aux chromosomes - aberrations chromosomiques ou formation de micronoyaux - peuvent être observés au microscope. Les dommages peuvent également être révélés en ajoutant un colorant aux cellules lors de la division cellulaire. L'exposition à un agent génotoxique peut alors être visualisée comme un échange accru du colorant entre les deux chromatides de chaque chromosome (échange de chromatides sœurs). Les aberrations chromosomiques sont liées à un risque accru de développer un cancer, mais la signification d'un taux accru d'échange de chromatides sœurs est moins claire.
Une évaluation plus sophistiquée de la génotoxicité est basée sur des mutations ponctuelles particulières dans les cellules somatiques, c'est-à-dire les globules blancs ou les cellules épithéliales obtenues à partir de la muqueuse buccale. Une mutation à un locus spécifique peut rendre les cellules capables de se développer dans une culture contenant un produit chimique autrement toxique (comme la 6-thioguanine). En variante, un produit génique spécifique peut être évalué (par exemple, des concentrations sériques ou tissulaires d'oncoprotéines codées par des oncogènes particuliers). De toute évidence, ces mutations reflètent le total des dommages génotoxiques subis et n'indiquent pas nécessairement quoi que ce soit sur l'exposition causale. Ces méthodes ne sont pas encore prêtes pour une utilisation pratique en médecine du travail, mais des progrès rapides dans cette ligne de recherche suggèrent que de telles méthodes deviendront disponibles d'ici quelques années.
Biomarqueurs de susceptibilité
Un marqueur de susceptibilité, qu'il soit héréditaire ou induit, est un indicateur que l'individu est particulièrement sensible à l'effet d'un xénobiotique ou aux effets d'un groupe de tels composés. La plus grande attention a été portée sur la susceptibilité génétique, bien que d'autres facteurs puissent être au moins aussi importants. L'hypersensibilité peut être due à un trait héréditaire, à la constitution de l'individu ou à des facteurs environnementaux.
La capacité à métaboliser certains produits chimiques est variable et déterminée génétiquement (voir « Déterminants génétiques de la réponse toxique »). Plusieurs enzymes pertinentes semblent être contrôlées par un seul gène. Par exemple, l'oxydation de produits chimiques étrangers est principalement réalisée par une famille d'enzymes appartenant à la famille P450. D'autres enzymes rendent les métabolites plus solubles dans l'eau par conjugaison (par exemple, N-acétyltransférase et μ-glutathion-S-transférase). L'activité de ces enzymes est contrôlée génétiquement et varie considérablement. Comme mentionné ci-dessus, l'activité peut être déterminée en administrant une petite dose d'un médicament, puis en déterminant la quantité du métabolite dans l'urine. Certains des gènes ont maintenant été caractérisés et des techniques sont disponibles pour déterminer le génotype. Des études importantes suggèrent qu'un risque de développer certaines formes de cancer est lié à la capacité de métaboliser des composés étrangers. De nombreuses questions restent encore en suspens, ce qui limite à ce jour l'utilisation de ces potentiels biomarqueurs de susceptibilité en santé au travail.
D'autres traits hérités, tels que l'alpha1-le déficit en antitrypsine ou déficit en glucose-6-phosphate déshydrogénase, entraînent également des mécanismes de défense déficients dans l'organisme, provoquant ainsi une hypersensibilité à certaines expositions.
La plupart des recherches liées à la susceptibilité ont porté sur la prédisposition génétique. D'autres facteurs jouent également un rôle et ont été en partie négligés. Par exemple, les personnes atteintes d'une maladie chronique peuvent être plus sensibles à une exposition professionnelle. De plus, si un processus pathologique ou une exposition antérieure à des produits chimiques toxiques a causé des dommages subcliniques aux organes, la capacité de résister à une nouvelle exposition toxique est susceptible d'être moindre. Des indicateurs biochimiques du fonctionnement des organes peuvent dans ce cas être utilisés comme biomarqueurs de susceptibilité. Le meilleur exemple concernant l'hypersensibilité concerne peut-être les réactions allergiques. Si un individu est devenu sensibilisé à une exposition particulière, des anticorps spécifiques peuvent être détectés dans le sérum. Même si l'individu n'a pas été sensibilisé, d'autres expositions actuelles ou passées peuvent augmenter le risque de développer un effet indésirable lié à une exposition professionnelle.
Un problème majeur est de déterminer l'effet conjoint des expositions mixtes au travail. De plus, les habitudes personnelles et la consommation de drogues peuvent entraîner une sensibilité accrue. Par exemple, la fumée de tabac contient généralement une quantité considérable de cadmium. Ainsi, avec une exposition professionnelle au cadmium, un gros fumeur qui a accumulé des quantités substantielles de ce métal dans le corps sera plus à risque de développer une maladie rénale liée au cadmium.
Application en santé au travail
Les biomarqueurs sont extrêmement utiles dans la recherche toxicologique et nombre d'entre eux peuvent s'appliquer à la surveillance biologique. Néanmoins, les limites doivent également être reconnues. De nombreux biomarqueurs n'ont jusqu'à présent été étudiés que sur des animaux de laboratoire. Les schémas toxicocinétiques chez d'autres espèces ne reflètent pas nécessairement la situation chez les êtres humains, et l'extrapolation peut nécessiter des études de confirmation chez des volontaires humains. Il faut également tenir compte des variations individuelles dues à des facteurs génétiques ou constitutionnels.
Dans certains cas, les biomarqueurs d'exposition peuvent ne pas être réalisables du tout (par exemple, pour les produits chimiques à courte durée de vie in vivo). D'autres produits chimiques peuvent être stockés dans des organes auxquels il n'est pas possible d'accéder par des procédures de routine, tels que le système nerveux, ou les affecter. La voie d'exposition peut également affecter le schéma de distribution et donc également la mesure du biomarqueur et son interprétation. Par exemple, une exposition directe du cerveau via le nerf olfactif est susceptible d'échapper à la détection par la mesure des biomarqueurs d'exposition. Quant aux biomarqueurs d'effet, beaucoup d'entre eux ne sont pas du tout spécifiques et le changement peut être dû à diverses causes, y compris des facteurs liés au mode de vie. Peut-être en particulier avec les biomarqueurs de sensibilité, l'interprétation doit être très prudente pour le moment, car de nombreuses incertitudes subsistent quant à l'importance globale pour la santé des génotypes individuels.
En santé au travail, le biomarqueur idéal doit répondre à plusieurs exigences. Tout d'abord, le prélèvement et l'analyse des échantillons doivent être simples et fiables. Pour une qualité analytique optimale, une normalisation est nécessaire, mais les exigences spécifiques varient considérablement. Les principaux domaines de préoccupation comprennent : la préparation de l'individu, la procédure d'échantillonnage et la manipulation des échantillons, et la procédure de mesure ; ce dernier englobe des facteurs techniques, tels que les procédures d'étalonnage et d'assurance qualité, et des facteurs liés à l'individu, tels que l'éducation et la formation des opérateurs.
Pour la documentation de la validité analytique et de la traçabilité, les matériaux de référence doivent être basés sur des matrices pertinentes et avec des concentrations appropriées de substances toxiques ou de métabolites pertinents à des niveaux appropriés. Pour que les biomarqueurs soient utilisés pour la surveillance biologique ou à des fins de diagnostic, les laboratoires responsables doivent disposer de procédures analytiques bien documentées avec des caractéristiques de performance définies et des enregistrements accessibles pour permettre la vérification des résultats. Dans le même temps, néanmoins, les aspects économiques de la caractérisation et de l'utilisation de matériaux de référence pour compléter les procédures d'assurance qualité en général doivent être pris en compte. Ainsi, la qualité réalisable des résultats et les utilisations qui en sont faites doivent être mises en balance avec les coûts supplémentaires de l'assurance qualité, y compris les matériaux de référence, la main-d'œuvre et l'instrumentation.
Une autre exigence est que le biomarqueur doit être spécifique, au moins dans les circonstances de l'étude, pour un type particulier d'exposition, avec une relation claire avec le degré d'exposition. Sinon, le résultat de la mesure du biomarqueur peut être trop difficile à interpréter. Pour une interprétation correcte du résultat de mesure d'un biomarqueur d'exposition, la validité diagnostique doit être connue (c'est-à-dire la traduction de la valeur du biomarqueur en ampleur des risques possibles pour la santé). Dans ce domaine, les métaux servent de paradigme pour la recherche de biomarqueurs. Des recherches récentes ont démontré la complexité et la subtilité des relations dose-réponse, avec des difficultés considérables pour identifier les niveaux sans effet et donc aussi pour définir les expositions tolérables. Cependant, ce type de recherche a également illustré les types d'enquête et le raffinement qui sont nécessaires pour découvrir les informations pertinentes. Pour la plupart des composés organiques, les associations quantitatives entre les expositions et les effets nocifs correspondants sur la santé ne sont pas encore disponibles ; dans de nombreux cas, même les principaux organes cibles ne sont pas connus avec certitude. De plus, l'évaluation des données de toxicité et des concentrations de biomarqueurs est souvent compliquée par l'exposition à des mélanges de substances, plutôt que par l'exposition à un seul composé à la fois.
Avant que le biomarqueur ne soit appliqué à des fins de santé au travail, certaines considérations supplémentaires sont nécessaires. Premièrement, le biomarqueur doit refléter uniquement un changement subclinique et réversible. Deuxièmement, étant donné que les résultats des biomarqueurs peuvent être interprétés en fonction des risques pour la santé, des efforts de prévention doivent être disponibles et doivent être considérés comme réalistes au cas où les données des biomarqueurs suggèrent la nécessité de réduire l'exposition. Troisièmement, l'utilisation pratique du biomarqueur doit être généralement considérée comme éthiquement acceptable.
Les mesures d'hygiène industrielle peuvent être comparées aux limites d'exposition applicables. De même, les résultats sur les biomarqueurs d'exposition ou les biomarqueurs d'effet peuvent être comparés à des limites d'action biologique, parfois appelées indices d'exposition biologique. Ces limites devraient être fondées sur les meilleurs conseils des cliniciens et des scientifiques des disciplines appropriées, et les administrateurs responsables en tant que « gestionnaires des risques » devraient alors tenir compte des facteurs éthiques, sociaux, culturels et économiques pertinents. La base scientifique devrait, si possible, inclure des relations dose-réponse complétées par des informations sur les variations de sensibilité au sein de la population à risque. Dans certains pays, les travailleurs et les membres du grand public sont impliqués dans le processus de normalisation et apportent une contribution importante, en particulier lorsque l'incertitude scientifique est considérable. L'une des principales incertitudes est de savoir comment définir un effet nocif sur la santé qui devrait être évité, par exemple, si la formation d'adduits en tant que biomarqueur d'exposition représente en soi un effet nocif (c'est-à-dire un biomarqueur d'effet) qui devrait être prévenu. Des questions difficiles sont susceptibles de se poser lorsqu'il s'agit de décider s'il est éthiquement défendable, pour un même composé, d'avoir des limites différentes pour l'exposition fortuite, d'une part, et l'exposition professionnelle, d'autre part.
Les informations générées par l'utilisation des biomarqueurs doivent généralement être transmises aux personnes examinées dans le cadre de la relation médecin-patient. Les préoccupations éthiques doivent notamment être prises en compte dans le cadre d'analyses de biomarqueurs très expérimentales qui ne peuvent actuellement être interprétées en détail en termes de risques réels pour la santé. Pour la population générale, par exemple, il existe actuellement peu d'orientations concernant l'interprétation des biomarqueurs d'exposition autres que la plombémie. La confiance dans les données générées est également importante (c'est-à-dire si un échantillonnage approprié a été effectué et si de bonnes procédures d'assurance qualité ont été utilisées dans le laboratoire concerné). Un domaine supplémentaire d'inquiétude particulière concerne l'hypersensibilité individuelle. Ces questions doivent être prises en compte lors de la restitution de l'étude.
Tous les secteurs de la société concernés par ou concernés par la réalisation d'une étude de biomarqueurs doivent être impliqués dans le processus de prise de décision sur la manière de traiter les informations générées par l'étude. Des procédures spécifiques pour prévenir ou surmonter les conflits éthiques inévitables doivent être développées dans les cadres juridiques et sociaux de la région ou du pays. Cependant, chaque situation représente un ensemble différent de questions et de pièges, et aucune procédure unique de participation du public ne peut être développée pour couvrir toutes les applications des biomarqueurs d'exposition.
Toxicologie des organes cibles
L'étude et la caractérisation des produits chimiques et autres agents pour leurs propriétés toxiques sont souvent entreprises sur la base d'organes et de systèmes d'organes spécifiques. Dans ce chapitre, deux cibles ont été sélectionnées pour une discussion approfondie : le système immunitaire et le gène. Ces exemples ont été choisis pour représenter un système d'organe cible complexe et une cible moléculaire dans les cellules. Pour une discussion plus complète de la toxicologie des organes cibles, le lecteur est renvoyé aux textes de toxicologie standard tels que Casarett et Doull, et Hayes. Le Programme international sur la sécurité chimique (IPCS) a également publié plusieurs documents de référence sur la toxicologie des organes cibles, par système d'organes.
Les études de toxicologie des organes cibles sont généralement entreprises sur la base d'informations indiquant le potentiel d'effets toxiques spécifiques d'une substance, soit à partir de données épidémiologiques, soit à partir d'études générales de toxicité aiguë ou chronique, soit sur la base de préoccupations particulières visant à protéger certaines fonctions d'organes, telles que que la reproduction ou le développement fœtal. Dans certains cas, des tests de toxicité pour des organes cibles spécifiques sont expressément mandatés par des autorités statutaires, tels que les tests de neurotoxicité en vertu de la loi américaine sur les pesticides (voir « L'approche des États-Unis pour l'évaluation des risques des toxiques pour la reproduction et des agents neurotoxiques », et les tests de mutagénicité en vertu de la loi japonaise sur les produits chimiques). Loi sur le contrôle des substances (voir « Principes d'identification des dangers : l'approche japonaise »).
Comme indiqué dans la section « Organe cible et effets critiques », l'identification d'un organe critique est basée sur la détection de l'organe ou du système d'organes qui réagit le premier négativement ou aux doses ou expositions les plus faibles. Ces informations sont ensuite utilisées pour concevoir des investigations toxicologiques spécifiques ou des tests de toxicité plus définis qui sont conçus pour élucider des indications plus sensibles d'intoxication dans l'organe cible. Les études de toxicologie des organes cibles peuvent également être utilisées pour déterminer les mécanismes d'action, utiles dans l'évaluation des risques (voir « L'approche des États-Unis pour l'évaluation des risques des toxiques pour la reproduction et des agents neurotoxiques »).
Méthodes d'études de toxicité pour les organes cibles
Les organes cibles peuvent être étudiés par exposition d'organismes intacts et analyse détaillée de la fonction et de l'histopathologie dans l'organe cible, ou par exposition in vitro de cellules, de tranches de tissus ou d'organes entiers maintenus pendant des périodes de courte ou de longue durée en culture (voir "Mécanismes de toxicologie : Introduction et concepts »). Dans certains cas, des tissus de sujets humains peuvent également être disponibles pour des études de toxicité sur des organes cibles, et ceux-ci peuvent fournir des opportunités pour valider les hypothèses d'extrapolation inter-espèces. Cependant, il faut garder à l'esprit que de telles études ne fournissent pas d'informations sur la toxicocinétique relative.
En général, les études de toxicité pour les organes cibles partagent les caractéristiques communes suivantes : examen histopathologique détaillé de l'organe cible, y compris examen post mortem, poids des tissus et examen des tissus fixés ; études biochimiques des voies critiques dans l'organe cible, telles que les systèmes enzymatiques importants ; des études fonctionnelles de la capacité de l'organe et des constituants cellulaires à remplir les fonctions métaboliques et autres attendues ; et l'analyse des biomarqueurs d'exposition et des effets précoces dans les cellules des organes cibles.
Une connaissance détaillée de la physiologie, de la biochimie et de la biologie moléculaire des organes cibles peut être intégrée aux études sur les organes cibles. Par exemple, étant donné que la synthèse et la sécrétion de protéines de faible poids moléculaire constituent un aspect important de la fonction rénale, les études de néphrotoxicité accordent souvent une attention particulière à ces paramètres (IPCS 1991). Étant donné que la communication de cellule à cellule est un processus fondamental du fonctionnement du système nerveux, les études d'organes cibles sur la neurotoxicité peuvent inclure des mesures neurochimiques et biophysiques détaillées de la synthèse, de l'absorption, du stockage, de la libération et de la liaison aux récepteurs des neurotransmetteurs, ainsi que des mesures électrophysiologiques des modifications de la membrane. potentiel associé à ces événements.
Un degré élevé d'accent est mis sur le développement de méthodes in vitro pour la toxicité des organes cibles, pour remplacer ou réduire l'utilisation d'animaux entiers. Des progrès substantiels dans ces méthodes ont été réalisés pour les substances toxiques pour la reproduction (Heindel et Chapin 1993).
En résumé, les études de toxicité pour les organes cibles sont généralement entreprises comme test d'ordre supérieur pour déterminer la toxicité. La sélection d'organes cibles spécifiques pour une évaluation plus approfondie dépend des résultats des tests de dépistage, tels que les tests aigus ou subchroniques utilisés par l'OCDE et l'Union européenne ; certains organes et systèmes d'organes cibles peuvent être candidats a priori à une enquête spéciale en raison du souci de prévenir certains types d'effets néfastes sur la santé.
Immunotoxicologie
Les fonctions du système immunitaire sont de protéger le corps contre les agents infectieux envahisseurs et d'assurer une surveillance immunitaire contre les cellules tumorales qui apparaissent. Il possède une première ligne de défense non spécifique qui peut initier elle-même des réactions effectrices, et une branche spécifique acquise, dans laquelle les lymphocytes et les anticorps portent la spécificité de reconnaissance et de réactivité ultérieure vis-à-vis de l'antigène.
L'immunotoxicologie a été définie comme « la discipline concernée par l'étude des événements pouvant entraîner des effets indésirables du fait de l'interaction des xénobiotiques avec le système immunitaire. Ces événements indésirables peuvent résulter (1) d'un effet direct et/ou indirect du xénobiotique (et/ou de son produit de biotransformation) sur le système immunitaire, ou (2) d'une réponse immunologique de l'hôte au composé et/ou son ou ses métabolites, ou des antigènes hôtes modifiés par le composé ou ses métabolites » (Berlin et al. 1987).
Lorsque le système immunitaire agit comme une cible passive d'agressions chimiques, il peut en résulter une diminution de la résistance aux infections et à certaines formes de néoplasie, ou une dérégulation/stimulation immunitaire pouvant exacerber les allergies ou l'auto-immunité. Dans le cas où le système immunitaire répond à la spécificité antigénique du xénobiotique ou de l'antigène hôte modifié par le composé, la toxicité peut se manifester sous forme d'allergies ou de maladies auto-immunes.
Des modèles animaux pour étudier la suppression immunitaire induite par des produits chimiques ont été développés, et un certain nombre de ces méthodes sont validées (Burleson, Munson et Dean 1995 ; IPCS 1996). À des fins de test, une approche à plusieurs niveaux est suivie pour effectuer une sélection adéquate parmi le nombre écrasant de tests disponibles. Généralement, l'objectif du premier niveau est d'identifier les immunotoxiques potentiels. Si une immunotoxicité potentielle est identifiée, un deuxième niveau de test est effectué pour confirmer et caractériser davantage les changements observés. Les enquêtes de troisième niveau comprennent des études spéciales sur le mécanisme d'action du composé. Plusieurs xénobiotiques ont été identifiés comme immunotoxiques provoquant une immunosuppression dans de telles études avec des animaux de laboratoire.
La base de données sur les perturbations de la fonction immunitaire chez les humains par les produits chimiques environnementaux est limitée (Descotes 1986; NRC Subcommittee on Immunotoxicology 1992). L'utilisation de marqueurs d'immunotoxicité a reçu peu d'attention dans les études cliniques et épidémiologiques pour étudier l'effet de ces produits chimiques sur la santé humaine. De telles études n'ont pas été réalisées fréquemment et leur interprétation ne permet souvent pas de tirer des conclusions univoques, en raison par exemple du caractère non contrôlé de l'exposition. Par conséquent, à l'heure actuelle, l'évaluation de l'immunotoxicité chez les rongeurs, avec extrapolation ultérieure à l'homme, constitue la base des décisions concernant les dangers et les risques.
Les réactions d'hypersensibilité, notamment l'asthme allergique et la dermatite de contact, sont d'importants problèmes de santé au travail dans les pays industrialisés (Vos, Younes et Smith 1995). Le phénomène de sensibilisation par contact a d'abord été étudié chez le cobaye (Andersen et Maibach 1985). Jusqu'à récemment, c'était l'espèce de choix pour les tests prédictifs. De nombreuses méthodes de test de cobaye sont disponibles, les plus fréquemment utilisées étant le test de maximisation du cobaye et le patch test occlus de Buehler. Les tests sur le cobaye et les nouvelles approches développées chez la souris, telles que les tests de gonflement des oreilles et le test des ganglions lymphatiques locaux, fournissent au toxicologue les outils nécessaires pour évaluer le risque de sensibilisation cutanée. La situation en matière de sensibilisation des voies respiratoires est très différente. Il n'existe pas encore de méthodes bien validées ou largement acceptées pour l'identification des allergènes respiratoires chimiques, bien que des progrès dans le développement de modèles animaux pour l'étude de l'allergie respiratoire chimique aient été réalisés chez le cobaye et la souris.
Les données humaines montrent que les agents chimiques, en particulier les médicaments, peuvent provoquer des maladies auto-immunes (Kammüller, Bloksma et Seinen 1989). Il existe un certain nombre de modèles animaux expérimentaux de maladies auto-immunes humaines. Ceux-ci comprennent à la fois une pathologie spontanée (par exemple le lupus érythémateux disséminé chez des souris noires de Nouvelle-Zélande) et des phénomènes auto-immuns induits par une immunisation expérimentale avec un auto-antigène à réaction croisée (par exemple l'arthrite induite par l'adjuvant H37Ra chez des rats de souche Lewis). Ces modèles sont appliqués dans l'évaluation préclinique des médicaments immunosuppresseurs. Très peu d'études ont abordé le potentiel de ces modèles pour évaluer si un xénobiotique exacerbe l'auto-immunité induite ou congénitale. Les modèles animaux qui conviennent pour étudier la capacité des produits chimiques à induire des maladies auto-immunes font pratiquement défaut. Un modèle qui est utilisé dans une mesure limitée est le test des ganglions lymphatiques poplités chez la souris. À l'instar de la situation chez l'homme, les facteurs génétiques jouent un rôle crucial dans le développement de maladies auto-immunes (MA) chez les animaux de laboratoire, ce qui limitera la valeur prédictive de tels tests.
Le système immunitaire
La fonction principale du système immunitaire est la défense contre les bactéries, les virus, les parasites, les champignons et les cellules néoplasiques. Ceci est réalisé par les actions de divers types de cellules et de leurs médiateurs solubles dans un concert finement réglé. La défense de l'hôte peut être grossièrement divisée en résistance non spécifique ou innée et en immunité spécifique ou acquise médiée par les lymphocytes (Roitt, Brostoff et Male 1989).
Les composants du système immunitaire sont présents dans tout le corps (Jones et al. 1990). Le compartiment lymphocytaire se trouve au sein des organes lymphoïdes (figure 1). La moelle osseuse et le thymus sont classés comme organes lymphoïdes primaires ou centraux ; les organes lymphoïdes secondaires ou périphériques comprennent les ganglions lymphatiques, la rate et le tissu lymphoïde le long des surfaces de sécrétion telles que les voies gastro-intestinales et respiratoires, le soi-disant tissu lymphoïde associé à la muqueuse (MALT). Environ la moitié des lymphocytes du corps se trouvent à tout moment dans le MALT. De plus, la peau est un organe important pour l'induction de réponses immunitaires aux antigènes présents sur la peau. Les cellules épidermiques de Langerhans qui ont une fonction de présentation d'antigène sont importantes dans ce processus.
Figure 1. Organes et tissus lymphoïdes primaires et secondaires
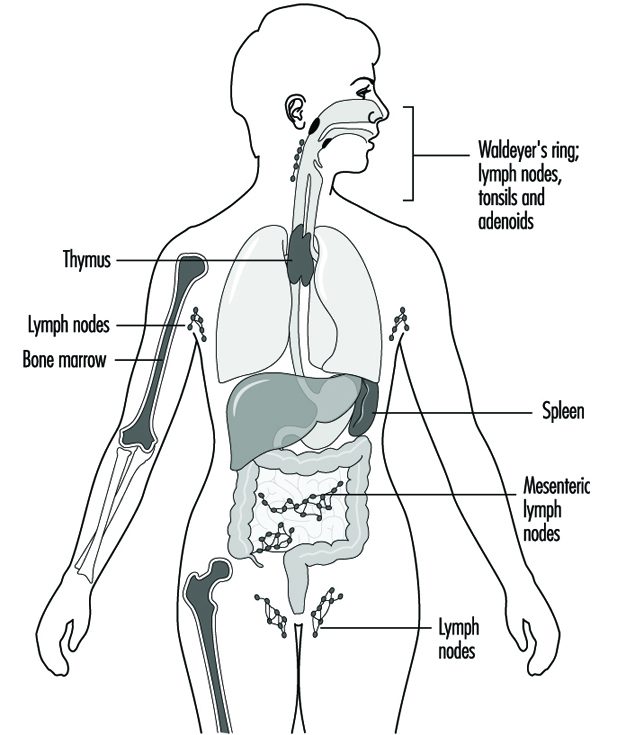
Les cellules phagocytaires de la lignée monocyte/macrophage, appelées système phagocytaire mononucléaire (MPS), sont présentes dans les organes lymphoïdes et également au niveau des sites extranodaux ; les phagocytes extranodaux comprennent les cellules de Kupffer dans le foie, les macrophages alvéolaires dans les poumons, les macrophages mésangiaux dans les reins et les cellules gliales dans le cerveau. Les leucocytes polymorphonucléaires (PMN) sont présents principalement dans le sang et la moelle osseuse, mais s'accumulent aux sites d'inflammation.
Défense non spécifique
Une première ligne de défense contre les micro-organismes est réalisée par une barrière physique et chimique, telle qu'au niveau de la peau, des voies respiratoires et du tube digestif. Cette barrière est aidée par des mécanismes de protection non spécifiques, notamment les cellules phagocytaires, telles que les macrophages et les leucocytes polymorphonucléaires, qui sont capables de tuer les agents pathogènes, et les cellules tueuses naturelles, qui peuvent lyser les cellules tumorales et les cellules infectées par des virus. Le système du complément et certains inhibiteurs microbiens (par exemple, le lysozyme) participent également à la réponse non spécifique.
Immunité spécifique
Après le contact initial de l'hôte avec l'agent pathogène, des réponses immunitaires spécifiques sont induites. La caractéristique de cette deuxième ligne de défense est la reconnaissance spécifique de déterminants, appelés antigènes ou épitopes, des agents pathogènes par des récepteurs à la surface cellulaire des lymphocytes B et T. Suite à l'interaction avec l'antigène spécifique, la cellule portant le récepteur est stimulée pour subir une prolifération et une différenciation, produisant un clone de cellules descendantes qui sont spécifiques de l'antigène déclenchant. Les réponses immunitaires spécifiques aident la défense non spécifique présentée aux agents pathogènes en stimulant l'efficacité des réponses non spécifiques. Une caractéristique fondamentale de l'immunité spécifique est que la mémoire se développe. Le contact secondaire avec le même antigène provoque une réponse plus rapide et plus vigoureuse mais bien régulée.
Le génome n'a pas la capacité de porter les codes d'un ensemble de récepteurs antigéniques suffisant pour reconnaître le nombre d'antigènes pouvant être rencontrés. Le répertoire de spécificité se développe par un processus de réarrangements de gènes. Il s'agit d'un processus aléatoire, au cours duquel différentes spécificités sont induites. Cela inclut des spécificités pour les composants autonomes, qui ne sont pas souhaitables. Un processus de sélection qui a lieu dans le thymus (cellules T), ou la moelle osseuse (cellules B) opère pour supprimer ces spécificités indésirables.
La fonction effectrice immunitaire normale et la régulation homéostatique de la réponse immunitaire dépendent d'une variété de produits solubles, connus collectivement sous le nom de cytokines, qui sont synthétisés et sécrétés par les lymphocytes et par d'autres types de cellules. Les cytokines ont des effets pléiotropes sur les réponses immunitaires et inflammatoires. La coopération entre différentes populations cellulaires est nécessaire pour la réponse immunitaire - la régulation des réponses d'anticorps, l'accumulation de cellules et de molécules immunitaires sur les sites inflammatoires, l'initiation de réponses de phase aiguë, le contrôle de la fonction cytotoxique des macrophages et de nombreux autres processus essentiels à la résistance de l'hôte . Celles-ci sont influencées par des cytokines agissant individuellement ou de concert, et dans de nombreux cas en dépendent.
Deux bras d'immunité spécifique sont reconnus - l'immunité humorale et l'immunité à médiation cellulaire ou cellulaire :
Immunité humorale. Dans le bras humoral, les lymphocytes B sont stimulés suite à la reconnaissance de l'antigène par les récepteurs de surface cellulaire. Les récepteurs antigéniques sur les lymphocytes B sont des immunoglobulines (Ig). Les lymphocytes B matures (cellules plasmatiques) déclenchent la production d'immunoglobulines spécifiques de l'antigène qui agissent comme des anticorps dans le sérum ou le long des surfaces muqueuses. Il existe cinq grandes classes d'immunoglobulines : (1) IgM, Ig pentamérique à capacité agglutinante optimale, qui est d'abord produite après stimulation antigénique ; (2) IgG, la principale Ig en circulation, qui peut passer le placenta ; (3) IgA, Ig sécrétoire pour la protection des surfaces muqueuses ; (4) IgE, Ig se fixant aux mastocytes ou aux granulocytes basophiles impliqués dans les réactions d'hypersensibilité immédiate et (5) IgD, dont la fonction principale est celle de récepteur sur les lymphocytes B.
Immunité à médiation cellulaire. Le bras cellulaire du système immunitaire spécifique est médié par les lymphocytes T. Ces cellules ont également des récepteurs antigéniques sur leurs membranes. Ils reconnaissent l'antigène s'il est présenté par des cellules présentatrices d'antigène dans le contexte des antigènes d'histocompatibilité. Par conséquent, ces cellules ont une restriction en plus de la spécificité antigénique. Les lymphocytes T fonctionnent comme des cellules auxiliaires pour diverses réponses immunitaires (y compris humorales), interviennent dans le recrutement de cellules inflammatoires et peuvent, en tant que lymphocytes T cytotoxiques, tuer les cellules cibles après la reconnaissance spécifique de l'antigène.
Mécanismes d'immunotoxicité
Immunosuppression
Une résistance efficace de l'hôte dépend de l'intégrité fonctionnelle du système immunitaire, qui à son tour exige que les cellules et molécules constitutives qui orchestrent les réponses immunitaires soient disponibles en nombre suffisant et sous une forme opérationnelle. Les immunodéficiences congénitales chez l'homme sont souvent caractérisées par des défauts dans certaines lignées de cellules souches, entraînant une production altérée ou absente de cellules immunitaires. Par analogie avec les maladies d'immunodéficience humaine congénitale et acquise, l'immunosuppression induite par des produits chimiques peut résulter simplement d'un nombre réduit de cellules fonctionnelles (IPCS 1996). L'absence ou le nombre réduit de lymphocytes peut avoir des effets plus ou moins profonds sur l'état immunitaire. Certains états d'immunodéficience et d'immunosuppression sévère, comme cela peut se produire lors d'une transplantation ou d'une thérapie cytostatique, ont été associés en particulier à des incidences accrues d'infections opportunistes et de certaines maladies néoplasiques. Les infections peuvent être bactériennes, virales, fongiques ou protozoaires, et le type prédominant d'infection dépend de l'immunodéficience associée. On peut s'attendre à ce que l'exposition à des produits chimiques environnementaux immunosuppresseurs entraîne des formes plus subtiles d'immunosuppression, qui peuvent être difficiles à détecter. Ceux-ci peuvent conduire, par exemple, à une incidence accrue d'infections telles que la grippe ou le rhume.
Compte tenu de la complexité du système immunitaire, avec la grande variété de cellules, de médiateurs et de fonctions qui forment un réseau compliqué et interactif, les composés immunotoxiques ont de nombreuses possibilités d'exercer un effet. Bien que la nature des lésions initiales induites par de nombreux produits chimiques immunotoxiques n'ait pas encore été élucidée, de plus en plus d'informations sont disponibles, principalement issues d'études sur des animaux de laboratoire, concernant les changements immunobiologiques qui entraînent une dépression de la fonction immunitaire (Dean et al. 1994) . Des effets toxiques peuvent se produire au niveau des fonctions critiques suivantes (et quelques exemples sont donnés de composés immunotoxiques affectant ces fonctions) :
- développement et expansion de différentes populations de cellules souches (le benzène exerce des effets immunotoxiques au niveau des cellules souches, provoquant une lymphocytopénie)
- prolifération de diverses cellules lymphoïdes et myéloïdes ainsi que des tissus de soutien dans lesquels ces cellules mûrissent et fonctionnent (les composés organostanniques immunotoxiques suppriment l'activité proliférative des lymphocytes dans le cortex thymique par cytotoxicité directe ; l'action thymotoxique du 2,3,7,8-tétrachloro -la dibenzo-p-dioxine (TCDD) et les composés apparentés sont probablement dus à une altération de la fonction des cellules épithéliales thymiques, plutôt qu'à une toxicité directe pour les thymocytes)
- l'absorption, le traitement et la présentation de l'antigène par les macrophages et d'autres cellules présentatrices d'antigène (l'une des cibles du 7,12-diméthylbenz(a)anthracène (DMBA) et du plomb est la présentation de l'antigène par les macrophages ; une cible du rayonnement ultraviolet est l'antigène- présentant la cellule de Langerhans)
- fonction régulatrice des cellules T-helper et T-suppressor (la fonction des cellules T-helper est altérée par les organostanniques, l'aldicarbe, les biphényles polychlorés (PCB), le TCDD et le DMBA ; la fonction des cellules T-suppressor est réduite par un traitement à faible dose de cyclophosphamide)
- production de diverses cytokines ou interleukines (le benzo(a)pyrène (BP) supprime la production d'interleukine-1 ; le rayonnement ultraviolet modifie la production de cytokines par les kératinocytes)
- la synthèse de diverses classes d'immunoglobulines IgM et IgG est supprimée après un traitement aux PCB et à l'oxyde de tributylétain (TBT) et augmentée après une exposition à l'hexachlorobenzène (HCB).
- régulation et activation du complément (affecté par TCDD)
- fonction des cellules T cytotoxiques (le 3-méthylcholanthrène (3-MC), le DMBA et le TCDD suppriment l'activité des cellules T cytotoxiques)
- fonction des cellules tueuses naturelles (NK) (l'activité NK pulmonaire est supprimée par l'ozone; l'activité NK splénique est altérée par le nickel)
- chimiotaxie des macrophages et des leucocytes polymorphonucléaires et fonctions cytotoxiques (l'ozone et le dioxyde d'azote altèrent l'activité phagocytaire des macrophages alvéolaires).
Allergie
Allergie peuvent être définis comme les effets néfastes sur la santé qui résultent de l'induction et du déclenchement de réponses immunitaires spécifiques. Lorsque des réactions d'hypersensibilité surviennent sans implication du système immunitaire, le terme pseudo-allergie est utilisé. Dans le contexte de l'immunotoxicologie, l'allergie résulte d'une réponse immunitaire spécifique aux produits chimiques et aux médicaments d'intérêt. La capacité d'un produit chimique à sensibiliser les individus est généralement liée à sa capacité à se lier de manière covalente aux protéines corporelles. Les réactions allergiques peuvent prendre diverses formes et celles-ci diffèrent en ce qui concerne à la fois les mécanismes immunologiques sous-jacents et la vitesse de la réaction. Quatre types principaux de réactions allergiques ont été reconnus : Réactions d'hypersensibilité de type I, qui sont provoquées par les anticorps IgE et où les symptômes se manifestent dans les minutes suivant l'exposition de l'individu sensibilisé. Les réactions d'hypersensibilité de type II résultent de l'endommagement ou de la destruction des cellules hôtes par les anticorps. Dans ce cas, les symptômes apparaissent en quelques heures. Les réactions d'hypersensibilité de type III, ou Arthus, sont également médiées par des anticorps, mais contre un antigène soluble, et résultent de l'action locale ou systémique de complexes immuns. Les réactions d'hypersensibilité de type IV, ou de type retardé, sont provoquées par les lymphocytes T et normalement les symptômes se développent 24 à 48 heures après l'exposition de l'individu sensibilisé.
Les deux types d'allergie chimique les plus pertinents pour la santé au travail sont la sensibilité de contact ou allergie cutanée et l'allergie des voies respiratoires.
Hypersensibilité de contact. Un grand nombre de produits chimiques sont capables de provoquer une sensibilisation cutanée. Suite à l'exposition topique d'un individu sensible à un allergène chimique, une réponse lymphocytaire T est induite dans les ganglions lymphatiques drainants. Dans la peau, l'allergène interagit directement ou indirectement avec les cellules épidermiques de Langerhans, qui transportent le produit chimique vers les ganglions lymphatiques et le présentent sous une forme immunogène aux lymphocytes T réactifs. Les lymphocytes T activés par les allergènes prolifèrent, entraînant une expansion clonale. L'individu est maintenant sensibilisé et répondra à une deuxième exposition cutanée au même produit chimique par une réponse immunitaire plus agressive, entraînant une dermatite de contact allergique. La réaction inflammatoire cutanée qui caractérise la dermatite allergique de contact est secondaire à la reconnaissance de l'allergène dans la peau par des lymphocytes T spécifiques. Ces lymphocytes s'activent, libèrent des cytokines et provoquent l'accumulation locale d'autres leucocytes mononucléaires. Les symptômes se développent environ 24 à 48 heures après l'exposition de l'individu sensibilisé, et la dermatite allergique de contact représente donc une forme d'hypersensibilité de type retardé. Les causes courantes de dermatite de contact allergique comprennent les produits chimiques organiques (tels que le 2,4-dinitrochlorobenzène), les métaux (tels que le nickel et le chrome) et les produits végétaux (tels que l'urushiol de l'herbe à puce).
Hypersensibilité respiratoire. L'hypersensibilité respiratoire est généralement considérée comme une réaction d'hypersensibilité de type I. Cependant, les réactions de phase tardive et les symptômes plus chroniques associés à l'asthme peuvent impliquer des processus immunitaires à médiation cellulaire (type IV). Les symptômes aigus associés à l'allergie respiratoire sont provoqués par des anticorps IgE dont la production est provoquée suite à l'exposition de l'individu sensible à l'allergène chimique inducteur. L'anticorps IgE se distribue de manière systémique et se lie, via des récepteurs membranaires, aux mastocytes qui se trouvent dans les tissus vascularisés, y compris les voies respiratoires. Suite à l'inhalation du même produit chimique, une réaction d'hypersensibilité respiratoire sera déclenchée. L'allergène s'associe aux protéines et se lie aux anticorps IgE liés aux mastocytes et les réticule. Cela provoque à son tour la dégranulation des mastocytes et la libération de médiateurs inflammatoires tels que l'histamine et les leucotriènes. De tels médiateurs provoquent une bronchoconstriction et une vasodilatation, entraînant des symptômes d'allergie respiratoire ; asthme et/ou rhinite. Les produits chimiques connus pour provoquer une hypersensibilité respiratoire chez l'homme comprennent les anhydrides acides (tels que l'anhydride trimellitique), certains diisocyanates (tels que le diisocyanate de toluène), les sels de platine et certains colorants réactifs. De plus, l'exposition chronique au béryllium est connue pour causer une maladie pulmonaire d'hypersensibilité.
auto-immunité
auto-immunité peut être défini comme la stimulation de réponses immunitaires spécifiques dirigées contre des antigènes endogènes du "soi". L'auto-immunité induite peut résulter soit d'altérations de l'équilibre des lymphocytes T régulateurs, soit de l'association d'un xénobiotique avec des composants tissulaires normaux de manière à les rendre immunogènes ("altéred self"). Les médicaments et les produits chimiques connus pour induire ou exacerber accidentellement des effets comme ceux de la maladie auto-immune (MA) chez les personnes sensibles sont des composés de faible poids moléculaire (poids moléculaire de 100 à 500) qui sont généralement considérés comme non immunogènes eux-mêmes. Le mécanisme de la MA par exposition chimique est pour la plupart inconnu. La maladie peut être produite directement au moyen d'anticorps circulants, indirectement par la formation de complexes immuns ou à la suite d'une immunité à médiation cellulaire, mais elle se produit probablement par une combinaison de mécanismes. La pathogénie est surtout connue dans les troubles hémolytiques immunitaires induits par les médicaments :
- Le médicament peut se fixer à la membrane des globules rouges et interagir avec un anticorps spécifique au médicament.
- Le médicament peut altérer la membrane des globules rouges de sorte que le système immunitaire considère la cellule comme étrangère.
- Le médicament et son anticorps spécifique forment des complexes immuns qui adhèrent à la membrane des globules rouges pour produire des lésions.
- La sensibilisation des globules rouges se produit en raison de la production d'auto-anticorps anti-globules rouges.
Une variété de produits chimiques et de médicaments, en particulier ces derniers, se sont avérés induire des réponses de type auto-immune (Kamüller, Bloksma et Seinen 1989). L'exposition professionnelle à des produits chimiques peut incidemment entraîner des syndromes de type MA. L'exposition au chlorure de vinyle monomère, au trichloroéthylène, au perchloroéthylène, aux résines époxy et à la poussière de silice peut induire des syndromes de type sclérodermique. Un syndrome similaire au lupus érythémateux disséminé (LED) a été décrit après exposition à l'hydrazine. L'exposition au diisocyanate de toluène a été associée à l'induction de purpura thrombocytopénique. Les métaux lourds tels que le mercure ont été impliqués dans certains cas de glomérulonéphrite à complexes immuns.
Évaluation des risques humains
L'évaluation de l'état immunitaire humain est effectuée principalement à l'aide de sang périphérique pour l'analyse de substances humorales telles que les immunoglobulines et le complément, et de leucocytes sanguins pour la composition de sous-ensembles et la fonctionnalité de sous-populations. Ces méthodes sont généralement les mêmes que celles utilisées pour étudier l'immunité humorale et à médiation cellulaire ainsi que la résistance non spécifique des patients suspectés d'immunodéficience congénitale. Pour les études épidémiologiques (par exemple, des populations professionnellement exposées), les paramètres doivent être sélectionnés sur la base de leur valeur prédictive dans les populations humaines, des modèles animaux validés et la biologie sous-jacente des marqueurs (voir tableau 1). La stratégie de dépistage des effets immunotoxiques après une exposition (accidentelle) à des polluants environnementaux ou à d'autres substances toxiques dépend beaucoup des circonstances, telles que le type d'immunodéficience à prévoir, le temps écoulé entre l'exposition et l'évaluation de l'état immunitaire, le degré d'exposition et le nombre d'individus exposés. Le processus d'évaluation du risque immunotoxique d'un xénobiotique particulier chez l'homme est extrêmement difficile et souvent impossible, en grande partie en raison de la présence de divers facteurs de confusion d'origine endogène ou exogène qui influencent la réponse des individus aux dommages toxiques. Cela est particulièrement vrai pour les études qui étudient le rôle de l'exposition chimique dans les maladies auto-immunes, où les facteurs génétiques jouent un rôle crucial.
Tableau 1. Classification des tests pour les marqueurs immunitaires
| Catégorie d'essai | Caractéristiques | Tests spécifiques |
| Général de base Doit être inclus avec les panneaux généraux |
Indicateurs de la santé générale et de l'état du système organique | Azote uréique sanguin, glycémie, etc. |
| Immunité basique Doit être inclus avec les panneaux généraux |
Indicateurs généraux de l'état immunitaire Coût relativement bas Les méthodes d'analyse sont standardisées entre les laboratoires Les résultats en dehors des plages de référence sont cliniquement interprétables |
Numérations sanguines complètes Taux sériques d'IgG, IgA, IgM Phénotypes de marqueurs de surface pour les principaux sous-ensembles de lymphocytes |
| Concentré/réflexe Doit être inclus lorsque cela est indiqué par des résultats cliniques, des expositions suspectées ou des résultats de tests antérieurs |
Indicateurs de fonctions/événements immunitaires spécifiques Le coût varie Les méthodes d'analyse sont standardisées entre les laboratoires Les résultats en dehors des plages de référence sont cliniquement interprétables |
Génotype d'histocompatibilité Anticorps contre les agents infectieux IgE sériques totales IgE spécifique de l'allergène Auto-anticorps Tests cutanés pour l'hypersensibilité Sursaut oxydatif des granulocytes Histopathologie (biopsie tissulaire) |
| Recherche Doit être inclus uniquement avec des populations témoins et une conception d'étude minutieuse |
Indicateurs de fonctions/événements immunitaires généraux ou spécifiques Le coût varie; souvent cher Les méthodes d'analyse ne sont généralement pas standardisées entre les laboratoires Les résultats en dehors des plages de référence ne sont souvent pas interprétables cliniquement |
Essais de stimulation in vitro Marqueurs de surface d'activation cellulaire Concentrations sériques de cytokines Essais de clonalité (anticorps, cellulaire, génétique) Tests de cytotoxicité |
Comme des données humaines adéquates sont rarement disponibles, l'évaluation du risque d'immunosuppression induite par des produits chimiques chez l'homme est dans la majorité des cas basée sur des études animales. L'identification des xénobiotiques immunotoxiques potentiels est entreprise principalement dans des études contrôlées chez les rongeurs. Les études d'exposition in vivo présentent, à cet égard, l'approche optimale pour estimer le potentiel immunotoxique d'un composé. Cela est dû à la nature multifactorielle et complexe du système immunitaire et des réponses immunitaires. Les études in vitro sont de plus en plus utiles pour élucider les mécanismes de l'immunotoxicité. De plus, en étudiant les effets du composé à l'aide de cellules d'origine animale et humaine, des données peuvent être générées pour la comparaison des espèces, qui peuvent être utilisées dans l'approche «parallélogramme» pour améliorer le processus d'évaluation des risques. Si des données sont disponibles pour les trois pierres angulaires du parallélogramme (animal in vivo, et animal in vitro et humain), il peut être plus facile de prédire le résultat à la pierre angulaire restante, c'est-à-dire le risque chez l'homme.
Lorsque l'évaluation du risque d'immunosuppression induite par des produits chimiques doit reposer uniquement sur des données provenant d'études animales, une approche peut être suivie dans l'extrapolation à l'homme en appliquant des facteurs d'incertitude à la dose sans effet nocif observé (NOAEL). Ce niveau peut être basé sur des paramètres déterminés dans des modèles pertinents, tels que des tests de résistance de l'hôte et une évaluation in vivo des réactions d'hypersensibilité et de la production d'anticorps. Idéalement, la pertinence de cette approche pour l'évaluation des risques nécessite une confirmation par des études chez l'homme. Ces études devraient combiner l'identification et la mesure de la substance toxique, les données épidémiologiques et les évaluations de l'état immunitaire.
Pour prédire l'hypersensibilité de contact, des modèles de cobayes sont disponibles et sont utilisés dans l'évaluation des risques depuis les années 1970. Bien que sensibles et reproductibles, ces tests ont des limites car ils dépendent d'une évaluation subjective ; cela peut être surmonté par des méthodes plus récentes et plus quantitatives développées chez la souris. Concernant l'hypersensibilité chimique induite par l'inhalation ou l'ingestion d'allergènes, des tests doivent être développés et évalués quant à leur valeur prédictive chez l'homme. Lorsqu'il s'agit de fixer des niveaux d'exposition professionnelle sûrs aux allergènes potentiels, il faut tenir compte de la nature biphasique de l'allergie : la phase de sensibilisation et la phase de déclenchement. La concentration requise pour déclencher une réaction allergique chez un individu précédemment sensibilisé est considérablement inférieure à la concentration nécessaire pour induire une sensibilisation chez l'individu immunologiquement naïf mais sensible.
Comme les modèles animaux pour prédire l'auto-immunité induite par des produits chimiques font pratiquement défaut, l'accent devrait être mis sur le développement de tels modèles. Pour le développement de tels modèles, nos connaissances sur l'auto-immunité induite par des produits chimiques chez l'homme doivent être avancées, y compris l'étude des marqueurs génétiques et du système immunitaire pour identifier les individus sensibles. Les humains qui sont exposés à des médicaments qui induisent l'auto-immunité offrent une telle opportunité.
Toxicologie génétique
La toxicologie génétique, par définition, est l'étude de la façon dont les agents chimiques ou physiques affectent le processus complexe de l'hérédité. Les produits chimiques génotoxiques sont définis comme des composés capables de modifier le matériel héréditaire des cellules vivantes. La probabilité qu'un produit chimique particulier cause des dommages génétiques dépend inévitablement de plusieurs variables, dont le niveau d'exposition de l'organisme au produit chimique, la distribution et la rétention du produit chimique une fois qu'il pénètre dans l'organisme, l'efficacité des systèmes d'activation métabolique et/ou de détoxification dans tissus cibles et la réactivité du produit chimique ou de ses métabolites avec les macromolécules critiques dans les cellules. La probabilité qu'un dommage génétique cause une maladie dépend en fin de compte de la nature du dommage, de la capacité de la cellule à réparer ou à amplifier le dommage génétique, de la possibilité d'exprimer toute altération induite et de la capacité de l'organisme à reconnaître et à supprimer la multiplication des cellules aberrantes.
Dans les organismes supérieurs, l'information héréditaire est organisée en chromosomes. Les chromosomes sont constitués de brins étroitement condensés d'ADN associé à des protéines. Au sein d'un même chromosome, chaque molécule d'ADN existe sous la forme d'une paire de longues chaînes non ramifiées de sous-unités nucléotidiques reliées entre elles par des liaisons phosphodiester qui relient le carbone 5 d'un fragment désoxyribose au carbone 3 du suivant (figure 1). De plus, l'une des quatre bases nucléotidiques différentes (adénine, cytosine, guanine ou thymine) est attachée à chaque sous-unité désoxyribose comme des perles sur une ficelle. En trois dimensions, chaque paire de brins d'ADN forme une double hélice avec toutes les bases orientées vers l'intérieur de la spirale. Au sein de l'hélice, chaque base est associée à sa base complémentaire sur le brin d'ADN opposé ; la liaison hydrogène dicte un appariement fort et non covalent de l'adénine avec la thymine et de la guanine avec la cytosine (figure 1). Étant donné que la séquence des bases nucléotidiques est complémentaire sur toute la longueur de la molécule d'ADN duplex, les deux brins portent essentiellement la même information génétique. En effet, lors de la réplication de l'ADN chaque brin sert de matrice pour la production d'un nouveau brin partenaire.
Figure 1. L'organisation (a) primaire, (b) secondaire et (c) tertiaire de l'information héréditaire humaine
 À l'aide d'ARN et d'un ensemble de protéines différentes, la cellule déchiffre finalement les informations codées par la séquence linéaire de bases dans des régions spécifiques de l'ADN (gènes) et produit des protéines essentielles à la survie cellulaire de base ainsi qu'à la croissance et à la différenciation normales. Essentiellement, les nucléotides fonctionnent comme un alphabet biologique utilisé pour coder les acides aminés, les éléments constitutifs des protéines.
À l'aide d'ARN et d'un ensemble de protéines différentes, la cellule déchiffre finalement les informations codées par la séquence linéaire de bases dans des régions spécifiques de l'ADN (gènes) et produit des protéines essentielles à la survie cellulaire de base ainsi qu'à la croissance et à la différenciation normales. Essentiellement, les nucléotides fonctionnent comme un alphabet biologique utilisé pour coder les acides aminés, les éléments constitutifs des protéines.
Lorsque des nucléotides incorrects sont insérés ou que des nucléotides sont perdus, ou lorsque des nucléotides inutiles sont ajoutés pendant la synthèse de l'ADN, l'erreur est appelée mutation. Il a été estimé que moins d'une mutation se produit pour 109 nucléotides incorporés lors de la réplication normale des cellules. Bien que les mutations ne soient pas nécessairement nocives, les altérations entraînant l'inactivation ou la surexpression de gènes importants peuvent entraîner divers troubles, notamment le cancer, des maladies héréditaires, des anomalies du développement, l'infertilité et la mort embryonnaire ou périnatale. Très rarement, une mutation peut entraîner une amélioration de la survie ; de tels événements sont à la base de la sélection naturelle.
Bien que certains produits chimiques réagissent directement avec l'ADN, la plupart nécessitent une activation métabolique. Dans ce dernier cas, les intermédiaires électrophiles tels que les époxydes ou les ions carbonium sont finalement responsables de l'induction de lésions sur une variété de sites nucléophiles au sein du matériel génétique (figure 2). Dans d'autres cas, la génotoxicité est médiée par des sous-produits de l'interaction du composé avec des lipides intracellulaires, des protéines ou de l'oxygène.
Figure 2. Bioactivation de : a) benzo(a)pyrène ; et b) N-nitrosodiméthylamine
En raison de leur abondance relative dans les cellules, les protéines sont la cible la plus fréquente des interactions toxiques. Cependant, la modification de l'ADN est plus préoccupante en raison du rôle central de cette molécule dans la régulation de la croissance et de la différenciation à travers plusieurs générations de cellules.
Au niveau moléculaire, les composés électrophiles ont tendance à attaquer l'oxygène et l'azote dans l'ADN. Les sites les plus susceptibles d'être modifiés sont illustrés à la figure 3. Bien que les oxygènes au sein des groupes phosphate dans le squelette de l'ADN soient également des cibles de modification chimique, on pense que les dommages aux bases sont biologiquement plus pertinents puisque ces groupes sont considérés comme le principal vecteur d'information. éléments de la molécule d'ADN.
Figure 3. Sites primaires de dommages à l'ADN induits chimiquement
Les composés qui contiennent une fraction électrophile exercent généralement une génotoxicité en produisant des mono-adduits dans l'ADN. De même, les composés qui contiennent deux fractions réactives ou plus peuvent réagir avec deux centres nucléophiles différents et produire ainsi des réticulations intra- ou inter-moléculaires dans le matériel génétique (figure 4). Les réticulations interbrin ADN-ADN et ADN-protéine peuvent être particulièrement cytotoxiques car elles peuvent former des blocs complets pour la réplication de l'ADN. Pour des raisons évidentes, la mort d'une cellule élimine la possibilité qu'elle soit mutée ou transformée de façon néoplasique. Les agents génotoxiques peuvent également agir en induisant des cassures dans le squelette phosphodiester, ou entre les bases et les sucres (produisant des sites abasiques) dans l'ADN. De telles cassures peuvent être le résultat direct de la réactivité chimique au niveau du site endommagé ou peuvent se produire pendant la réparation de l'un des types de lésions de l'ADN susmentionnés.
Figure 4. Différents types de dommages au complexe protéine-ADN
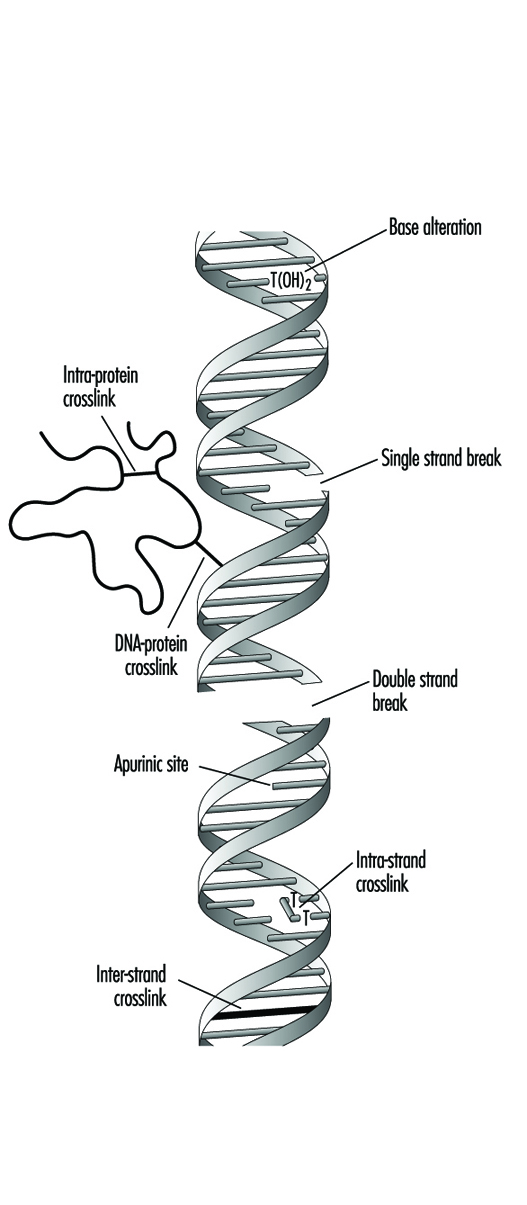
Au cours des trente à quarante dernières années, diverses techniques ont été développées pour surveiller le type de dommages génétiques induits par divers produits chimiques. Ces tests sont décrits en détail ailleurs dans ce chapitre et Encyclopédie.
Une mauvaise réplication de « microlésions » telles que des mono-adduits, des sites abasiques ou des cassures simple brin peut finalement entraîner des substitutions de paires de bases de nucléotides, ou l'insertion ou la suppression de courts fragments de polynucléotides dans l'ADN chromosomique. En revanche, les «macrolésions», telles que les adduits volumineux, les réticulations ou les cassures double brin, peuvent déclencher le gain, la perte ou le réarrangement de morceaux de chromosomes relativement volumineux. Dans tous les cas, les conséquences peuvent être dévastatrices pour l'organisme puisque chacun de ces événements peut entraîner la mort cellulaire, la perte de fonction ou la transformation maligne des cellules. La manière exacte dont les dommages à l'ADN causent le cancer est en grande partie inconnue. On pense actuellement que le processus peut impliquer une activation inappropriée de proto-oncogènes tels que monc ainsi que ras, et/ou l'inactivation de gènes suppresseurs de tumeurs récemment identifiés tels que p53. L'expression anormale de l'un ou l'autre type de gène abroge les mécanismes cellulaires normaux pour contrôler la prolifération et/ou la différenciation cellulaire.
La prépondérance des preuves expérimentales indique que le développement d'un cancer suite à une exposition à des composés électrophiles est un événement relativement rare. Cela peut s'expliquer, en partie, par la capacité intrinsèque de la cellule à reconnaître et à réparer l'ADN endommagé ou par l'incapacité des cellules dont l'ADN est endommagé à survivre. Pendant la réparation, la base endommagée, le nucléotide ou le court tronçon de nucléotides entourant le site endommagé est retiré et (en utilisant le brin opposé comme modèle) un nouveau morceau d'ADN est synthétisé et épissé en place. Pour être efficace, la réparation de l'ADN doit se produire avec une grande précision avant la division cellulaire, avant les opportunités de propagation de la mutation.
Des études cliniques ont montré que les personnes présentant des défauts héréditaires dans la capacité de réparer l'ADN endommagé développent fréquemment un cancer et/ou des anomalies du développement à un âge précoce (tableau 1). De tels exemples fournissent des preuves solides reliant l'accumulation de dommages à l'ADN à la maladie humaine. De même, les agents qui favorisent la prolifération cellulaire (tels que l'acétate de tétradécanoylphorbol) améliorent souvent la carcinogenèse. Pour ces composés, la probabilité accrue de transformation néoplasique peut être une conséquence directe d'une diminution du temps disponible pour que la cellule effectue une réparation adéquate de l'ADN.
Tableau 1. Troubles héréditaires prédisposés au cancer qui semblent impliquer des défauts de réparation de l'ADN
| Syndrome | Symptômes | Phénotype cellulaire |
| Ataxie télangiectasie | Détérioration neurologique Immunodéficience Incidence élevée de lymphome |
Hypersensibilité aux rayonnements ionisants et à certains agents alkylants. Réplication dérégulée de l'ADN endommagé (peut indiquer un temps raccourci pour la réparation de l'ADN) |
| Syndrome de Bloom | Anomalies du développement Lésions sur la peau exposée Incidence élevée de tumeurs du système immunitaire et du tractus gastro-intestinal |
Fréquence élevée des aberrations chromosomiques Ligature défectueuse des cassures associées à la réparation de l'ADN |
| L'anémie de Fanconi | Retard de croissance Incidence élevée de leucémie |
Hypersensibilité aux agents de réticulation Fréquence élevée des aberrations chromosomiques Réparation défectueuse des liaisons croisées dans l'ADN |
| Cancer du côlon héréditaire sans polypose | Forte incidence du cancer du côlon | Défaut dans la réparation des mésappariements d'ADN (lorsque l'insertion d'un mauvais nucléotide se produit pendant la réplication) |
| Xéroderma pigmentosum | Incidence élevée d'épithéliome sur les zones exposées de la peau Atteinte neurologique (dans de nombreux cas) |
Hypersensibilité aux rayons UV et à de nombreux cancérigènes chimiques Défauts de réparation par excision et/ou de réplication de l'ADN endommagé |
Les premières théories sur la façon dont les produits chimiques interagissent avec l'ADN remontent aux études menées lors du développement du gaz moutarde utilisé dans la guerre. Une meilleure compréhension est née des efforts visant à identifier des agents anticancéreux qui arrêteraient sélectivement la réplication des cellules tumorales à division rapide. L'inquiétude croissante du public concernant les dangers dans notre environnement a incité des recherches supplémentaires sur les mécanismes et les conséquences de l'interaction chimique avec le matériel génétique. Des exemples de divers types de produits chimiques qui exercent une génotoxicité sont présentés dans le tableau 2.
Tableau 2. Exemples de produits chimiques qui présentent une génotoxicité dans les cellules humaines
| Classe de produit chimique | Exemple | Source d'exposition | Lésion génotoxique probable |
| Aflatoxines | Aflatoxine B1 | Nourriture contaminée | Adduits volumineux à l'ADN |
| Amines aromatiques | 2-Acétylaminofluorène | Environnement | Adduits volumineux à l'ADN |
| Quinones d'aziridine | Mitomycine c | Chimiothérapie anticancéreuse | Mono-adduits, réticulations interbrins et cassures simple brin dans l'ADN. |
| Hydrocarbures chlorés | Chlorure de vinyle | Environnement | Mono-adduits dans l'ADN |
| Métaux et composés métalliques | Cisplatine | Chimiothérapie anticancéreuse | Crosslinks intra- et inter-brins dans l'ADN |
| Composés de nickel | Environnement | Mono-adduits et cassures simple brin dans l'ADN | |
| Moutardes à l'azote | Cyclophosphamide | Chimiothérapie anticancéreuse | Mono-adduits et réticulations interbrins dans l'ADN |
| Nitrosamines | N-Nitrosodiméthylamine | Nourriture contaminée | Mono-adduits dans l'ADN |
| Hydrocarbures aromatiques polycycliques | Benzo (a) pyrène | Environnement | Adduits volumineux à l'ADN |
Lésion cellulaire et mort cellulaire
Pratiquement toute la médecine est consacrée soit à prévenir la mort cellulaire, dans des maladies telles que l'infarctus du myocarde, les accidents vasculaires cérébraux, les traumatismes et les chocs, soit à la provoquer, comme dans le cas des maladies infectieuses et du cancer. Il est donc essentiel d'en comprendre la nature et les mécanismes impliqués. La mort cellulaire a été classée comme « accidentelle », c'est-à-dire causée par des agents toxiques, l'ischémie, etc., ou « programmée », comme cela se produit au cours du développement embryologique, y compris la formation des doigts et la résorption de la queue du têtard.
Les lésions cellulaires et la mort cellulaire sont donc importantes à la fois en physiologie et en physiopathologie. La mort cellulaire physiologique est extrêmement importante au cours de l'embryogenèse et du développement embryonnaire. L'étude de la mort cellulaire au cours du développement a conduit à des informations importantes et nouvelles sur la génétique moléculaire impliquée, notamment à travers l'étude du développement chez les animaux invertébrés. Chez ces animaux, la localisation précise et la signification des cellules destinées à subir la mort cellulaire ont été soigneusement étudiées et, grâce à l'utilisation des techniques classiques de mutagénèse, plusieurs gènes impliqués ont maintenant été identifiés. Dans les organes adultes, l'équilibre entre la mort cellulaire et la prolifération cellulaire contrôle la taille de l'organe. Dans certains organes, comme la peau et l'intestin, il y a un renouvellement continu des cellules. Dans la peau, par exemple, les cellules se différencient lorsqu'elles atteignent la surface, et subissent finalement une différenciation terminale et la mort cellulaire au fur et à mesure que la kératinisation se poursuit avec la formation d'enveloppes réticulées.
De nombreuses classes de produits chimiques toxiques sont capables d'induire des lésions cellulaires aiguës suivies de la mort. Ceux-ci comprennent l'anoxie et l'ischémie et leurs analogues chimiques tels que le cyanure de potassium ; les cancérigènes chimiques, qui forment des électrophiles qui se lient de manière covalente aux protéines des acides nucléiques ; des produits chimiques oxydants, entraînant la formation de radicaux libres et des lésions oxydantes ; activation du complément ; et une variété d'ionophores de calcium. La mort cellulaire est également une composante importante de la carcinogenèse chimique; de nombreux carcinogènes chimiques complets, à des doses cancérigènes, produisent une nécrose et une inflammation aiguës suivies d'une régénération et d'une prénéoplasie.
Définitions
Lésion cellulaire
Une lésion cellulaire est définie comme un événement ou un stimulus, tel qu'un produit chimique toxique, qui perturbe l'homéostasie normale de la cellule, provoquant ainsi un certain nombre d'événements (figure 1). Les principales cibles des lésions mortelles illustrées sont l'inhibition de la synthèse d'ATP, la perturbation de l'intégrité de la membrane plasmique ou le retrait des facteurs de croissance essentiels.
Les blessures mortelles entraînent la mort d'une cellule après une période de temps variable, en fonction de la température, du type de cellule et du stimulus ; ou ils peuvent être sublétaux ou chroniques, c'est-à-dire que la lésion entraîne une altération de l'homéostasie qui, bien qu'anormale, n'entraîne pas la mort cellulaire (Trump et Arstila 1971 ; Trump et Berezesky 1992 ; Trump et Berezesky 1995 ; Trump, Berezesky et Osornio-Vargas 1981). Dans le cas d'une blessure mortelle, il y a une phase avant le moment de la mort cellulaire
pendant ce temps, la cellule récupérera; cependant, après un moment donné (le "point de non-retour" ou le point de mort cellulaire), l'élimination de la blessure n'entraîne pas de récupération, mais la cellule subit une dégradation et une hydrolyse, atteignant finalement un équilibre physico-chimique avec le environnement. C'est la phase dite de nécrose. Au cours de la phase prélétale, plusieurs types principaux de changements se produisent, selon la cellule et le type de blessure. Celles-ci sont connues sous le nom d'apoptose et d'oncose.
L'apoptose
L'apoptose est dérivé des mots grecs apo, c'est-à-dire loin de, et ptosis, signifiant tomber. Le terme s'éloigner de vient du fait que, lors de ce type de changement prélétal, les cellules se rétractent et subissent un important bourgeonnement en périphérie. Les bulles se détachent alors et flottent. L'apoptose se produit dans une variété de types de cellules suite à divers types de lésions toxiques (Wyllie, Kerr et Currie 1980). Il est particulièrement important dans les lymphocytes, où il est le mécanisme prédominant de renouvellement des clones de lymphocytes. Les fragments résultants donnent les corps basophiles observés dans les macrophages des ganglions lymphatiques. Dans d'autres organes, l'apoptose se produit typiquement dans des cellules individuelles qui sont rapidement éliminées avant et après la mort par phagocytose des fragments par des cellules parenchymateuses adjacentes ou par des macrophages. L'apoptose survenant dans des cellules individuelles avec phagocytose ultérieure n'entraîne généralement pas d'inflammation. Avant la mort, les cellules apoptotiques présentent un cytosol très dense avec des mitochondries normales ou condensées. Le réticulum endoplasmique (RE) est normal ou peu dilaté. La chromatine nucléaire est nettement agglutinée le long de l'enveloppe nucléaire et autour du nucléole. Le contour nucléaire est également irrégulier et une fragmentation nucléaire se produit. La condensation de la chromatine est associée à la fragmentation de l'ADN qui, dans de nombreux cas, se produit entre les nucléosomes, donnant un aspect caractéristique en échelle lors de l'électrophorèse.
En apoptose, augmentation de [Ca2+]i peut stimuler K+ efflux entraînant un rétrécissement cellulaire, ce qui nécessite probablement de l'ATP. Les blessures qui inhibent totalement la synthèse d'ATP sont donc plus susceptibles d'entraîner l'apoptose. Une augmentation soutenue de [Ca2+]i a un certain nombre d'effets délétères, y compris l'activation des protéases, des endonucléases et des phospholipases. L'activation de l'endonucléase entraîne des ruptures de brins d'ADN simples et doubles qui, à leur tour, stimulent des niveaux accrus de p53 et de ribosylation poly-ADP, et de protéines nucléaires essentielles à la réparation de l'ADN. L'activation des protéases modifie un certain nombre de substrats, y compris l'actine et les protéines apparentées, conduisant à la formation de bulles. Un autre substrat important est la poly(ADP-ribose) polymérase (PARP), qui inhibe la réparation de l'ADN. Augmentation de [Ca2+]i est également associée à l'activation d'un certain nombre de protéines kinases, telles que la MAP kinase, la calmoduline kinase et autres. Ces kinases sont impliquées dans l'activation des facteurs de transcription qui initient la transcription des gènes précoces immédiats, par exemple, c-fos, c-jun et c-myc, et dans l'activation de la phospholipase A2 ce qui se traduit par une perméabilisation de la membrane plasmique et des membranes intracellulaires telles que la membrane interne des mitochondries.
Oncose
Oncose, dérivé du mot grec Est-ce que s, gonfler, est ainsi nommé parce que dans ce type de changement prélétal, la cellule commence à gonfler presque immédiatement après la blessure (Majno et Joris 1995). La raison du gonflement est une augmentation des cations dans l'eau à l'intérieur de la cellule. Le principal cation responsable est le sodium, qui est normalement régulé pour maintenir le volume cellulaire. Cependant, en l'absence d'ATP ou si la Na-ATPase du plasmalemme est inhibée, le contrôle du volume est perdu à cause des protéines intracellulaires et le sodium dans l'eau continue d'augmenter. Parmi les événements précoces de l'oncose sont donc augmentés [Na+]i ce qui conduit à un gonflement cellulaire et à une augmentation de [Ca2+]i résultant soit de l'influx de l'espace extracellulaire, soit de la libération des réserves intracellulaires. Il en résulte un gonflement du cytosol, un gonflement du réticulum endoplasmique et de l'appareil de Golgi, et la formation de bulles aqueuses autour de la surface cellulaire. Les mitochondries subissent initialement une condensation, mais plus tard, elles présentent également un gonflement de grande amplitude en raison de dommages à la membrane mitochondriale interne. Dans ce type de changement prélétal, la chromatine subit une condensation et finalement une dégradation ; cependant, le modèle d'échelle caractéristique de l'apoptose n'est pas observé.
Nécrose
La nécrose fait référence à la série de changements qui se produisent après la mort cellulaire lorsque la cellule est convertie en débris qui sont généralement éliminés par la réponse inflammatoire. Deux types peuvent être distingués : la nécrose oncotique et la nécrose apoptotique. La nécrose oncotique survient généralement dans de grandes zones, par exemple, dans un infarctus du myocarde ou régionalement dans un organe après une toxicité chimique, comme le tubule rénal proximal après administration de HgCl2. De larges zones d'un organe sont atteintes et les cellules nécrotiques provoquent rapidement une réaction inflammatoire, d'abord aiguë puis chronique. En cas de survie de l'organisme, dans de nombreux organes, la nécrose est suivie d'une élimination des cellules mortes et d'une régénération, par exemple dans le foie ou les reins suite à une toxicité chimique. En revanche, la nécrose apoptotique se produit généralement sur une seule cellule et les débris nécrotiques se forment dans les phagocytes des macrophages ou des cellules parenchymateuses adjacentes. Les premières caractéristiques des cellules nécrotiques comprennent des interruptions dans la continuité de la membrane plasmique et l'apparition de densités floconneuses, représentant des protéines dénaturées au sein de la matrice mitochondriale. Dans certaines formes de lésions qui n'interfèrent pas initialement avec l'accumulation de calcium mitochondrial, des dépôts de phosphate de calcium peuvent être observés dans les mitochondries. D'autres systèmes membranaires se fragmentent de la même manière, tels que le RE, les lysosomes et l'appareil de Golgi. En fin de compte, la chromatine nucléaire subit une lyse, résultant de l'attaque par les hydrolases lysosomales. Après la mort cellulaire, les hydrolases lysosomales jouent un rôle important dans l'élimination des débris avec les cathepsines, les nucléolases et les lipases, car celles-ci ont un pH acide optimal et peuvent survivre au faible pH des cellules nécrotiques tandis que d'autres enzymes cellulaires sont dénaturées et inactivées.
Mécanismes
Stimulus initial
Dans le cas de lésions mortelles, les interactions initiales les plus courantes entraînant une lésion entraînant la mort cellulaire sont l'interférence avec le métabolisme énergétique, comme l'anoxie, l'ischémie ou les inhibiteurs de la respiration, et la glycolyse comme le cyanure de potassium, le monoxyde de carbone, l'iodo-acétate et bientôt. Comme mentionné ci-dessus, des doses élevées de composés qui inhibent le métabolisme énergétique entraînent généralement une oncose. L'autre type courant de lésion initiale entraînant une mort cellulaire aiguë est la modification de la fonction de la membrane plasmique (Trump et Arstila 1971 ; Trump, Berezesky et Osornio-Vargas 1981). Cela peut être soit des dommages directs et une perméabilisation, comme dans le cas d'un traumatisme ou de l'activation du complexe C5b-C9 du complément, des dommages mécaniques à la membrane cellulaire ou une inhibition du sodium-potassium (Na+-K+) pompe avec des glycosides tels que l'ouabaïne. Les ionophores calciques tels que l'ionomycine ou A23187, qui transportent rapidement [Ca2+] vers le bas du gradient dans la cellule, provoquent également des blessures mortelles aiguës. Dans certains cas, le schéma du changement prélétal est l'apoptose ; dans d'autres, c'est une oncose.
Voies de signalisation
Avec de nombreux types de lésions, la respiration mitochondriale et la phosphorylation oxydative sont rapidement affectées. Dans certaines cellules, cela stimule la glycolyse anaérobie, qui est capable de maintenir l'ATP, mais avec de nombreuses blessures, cela est inhibé. Le manque d'ATP entraîne une incapacité à dynamiser un certain nombre de processus homéostatiques importants, en particulier le contrôle de l'homéostasie des ions intracellulaires (Trump et Berezesky 1992 ; Trump, Berezesky et Osornio-Vargas 1981). Il en résulte une augmentation rapide de [Ca2+]i, et augmenté [Na+] et [Cl-] entraîne un gonflement des cellules. Augmentation de [Ca2+]i entraîner l'activation d'un certain nombre d'autres mécanismes de signalisation discutés ci-dessous, y compris une série de kinases, ce qui peut entraîner une augmentation immédiate de la transcription précoce des gènes. Augmentation de [Ca2+]i modifie également la fonction cytosquelettique, entraînant en partie la formation de bulles et l'activation des endonucléases, des protéases et des phospholipases. Ceux-ci semblent déclencher bon nombre des effets importants discutés ci-dessus, tels que les dommages à la membrane par l'activation de la protéase et de la lipase, la dégradation directe de l'ADN à partir de l'activation de l'endonucléase et l'activation de kinases telles que la MAP kinase et la calmoduline kinase, qui agissent comme facteurs de transcription.
Grâce à un travail approfondi sur le développement chez les invertébrés C. elegans ainsi que Drosophila, ainsi que des cellules humaines et animales, une série de gènes pro-mort ont été identifiés. Certains de ces gènes d'invertébrés se sont avérés avoir des homologues de mammifères. Par exemple, le gène ced-3, essentiel à la mort cellulaire programmée chez C. elegans, a une activité protéase et une forte homologie avec l'enzyme de conversion de l'interleukine de mammifère (ICE). Un gène étroitement apparenté appelé apopain ou prICE a récemment été identifié avec une homologie encore plus étroite (Nicholson et al. 1995). Dans Drosophila, le gène reaper semble être impliqué dans un signal qui conduit à la mort cellulaire programmée. D'autres gènes pro-mort comprennent la protéine membranaire Fas et l'important gène suppresseur de tumeur, p53, qui est largement conservé. p53 est induit au niveau protéique suite à des dommages à l'ADN et, lorsqu'il est phosphorylé, agit comme un facteur de transcription pour d'autres gènes tels que gadd45 et waf-1, qui sont impliqués dans la signalisation de la mort cellulaire. D'autres gènes précoces immédiats tels que c-fos, c-jun et c-myc semblent également être impliqués dans certains systèmes.
En même temps, il existe des gènes anti-mort qui semblent contrecarrer les gènes pro-mort. Le premier d'entre eux à être identifié était ced-9 de C. elegans, qui est homologue à bcl-2 chez l'homme. Ces gènes agissent d'une manière encore inconnue pour empêcher la destruction des cellules par des toxines génétiques ou chimiques. Certaines preuves récentes indiquent que bcl-2 peut agir comme un antioxydant. Actuellement, de nombreux efforts sont en cours pour développer une compréhension des gènes impliqués et pour développer des moyens d'activer ou d'inhiber ces gènes, selon la situation.
Introduction et notions
La toxicologie mécaniste est l'étude de la façon dont les agents chimiques ou physiques interagissent avec les organismes vivants pour provoquer une toxicité. La connaissance du mécanisme de toxicité d'une substance améliore la capacité à prévenir la toxicité et à concevoir des produits chimiques plus souhaitables ; elle constitue la base de la thérapie en cas de surexposition et permet souvent une meilleure compréhension des processus biologiques fondamentaux. Aux fins de ce Encyclopédie l'accent sera mis sur les animaux pour prédire la toxicité humaine. Les différents domaines de la toxicologie comprennent la toxicologie mécaniste, descriptive, réglementaire, médico-légale et environnementale (Klaassen, Amdur et Doull 1991). Tous ces éléments bénéficient de la compréhension des mécanismes fondamentaux de la toxicité.
Pourquoi comprendre les mécanismes de toxicité ?
Comprendre le mécanisme par lequel une substance provoque une toxicité améliore différents domaines de la toxicologie de différentes manières. La compréhension mécaniste aide le régulateur gouvernemental à établir des limites de sécurité juridiquement contraignantes pour l'exposition humaine. Il aide les toxicologues à recommander des plans d'action concernant le nettoyage ou l'assainissement des sites contaminés et, avec les propriétés physiques et chimiques de la substance ou du mélange, peut être utilisé pour sélectionner le degré d'équipement de protection requis. Les connaissances mécanistes sont également utiles pour former la base de la thérapie et de la conception de nouveaux médicaments pour le traitement des maladies humaines. Pour le toxicologue médico-légal, le mécanisme de la toxicité donne souvent un aperçu de la façon dont un agent chimique ou physique peut causer la mort ou une incapacité.
Si le mécanisme de la toxicité est compris, la toxicologie descriptive devient utile pour prédire les effets toxiques des produits chimiques apparentés. Il est important de comprendre, cependant, qu'un manque d'informations mécanistes ne dissuade pas les professionnels de la santé de protéger la santé humaine. Des décisions prudentes basées sur des études animales et l'expérience humaine sont utilisées pour établir des niveaux d'exposition sûrs. Traditionnellement, une marge de sécurité était établie en utilisant le « niveau sans effet nocif » ou le « niveau le plus faible avec effet nocif » provenant d'études sur des animaux (en utilisant des modèles d'exposition répétée) et en divisant ce niveau par un facteur de 100 pour l'exposition professionnelle ou de 1,000 XNUMX pour l'exposition professionnelle. autre exposition environnementale humaine. Le succès de ce processus est évident d'après les quelques incidents d'effets néfastes sur la santé attribués à l'exposition chimique chez les travailleurs pour lesquels des limites d'exposition appropriées avaient été fixées et respectées dans le passé. De plus, la durée de vie humaine continue d'augmenter, tout comme la qualité de vie. Dans l'ensemble, l'utilisation des données de toxicité a conduit à un contrôle réglementaire et volontaire efficace. Une connaissance détaillée des mécanismes toxiques améliorera la prévisibilité des nouveaux modèles de risque en cours d'élaboration et se traduira par une amélioration continue.
La compréhension des mécanismes environnementaux est complexe et suppose une connaissance des perturbations et de l'homéostasie (équilibre) des écosystèmes. Bien que cela ne soit pas abordé dans cet article, une meilleure compréhension des mécanismes toxiques et de leurs conséquences ultimes dans un écosystème aiderait les scientifiques à prendre des décisions prudentes concernant la manipulation des déchets municipaux et industriels. La gestion des déchets est un domaine de recherche en plein essor et continuera d'être très importante à l'avenir.
Techniques d'étude des mécanismes de toxicité
La majorité des études mécanistes débutent par une étude toxicologique descriptive chez l'animal ou des observations cliniques chez l'homme. Idéalement, les études animales comprennent des observations comportementales et cliniques minutieuses, un examen biochimique minutieux des éléments du sang et de l'urine à la recherche de signes de fonctionnement indésirable des principaux systèmes biologiques de l'organisme, et une évaluation post-mortem de tous les systèmes d'organes par examen microscopique pour vérifier blessure (voir les directives d'essai de l'OCDE ; les directives de la CE sur l'évaluation des produits chimiques ; les règles d'essai de l'EPA des États-Unis ; la réglementation japonaise sur les produits chimiques). Ceci est analogue à un examen physique humain approfondi qui aurait lieu dans un hôpital sur une période de deux à trois jours, à l'exception de l'autopsie.
Comprendre les mécanismes de la toxicité est l'art et la science de l'observation, la créativité dans la sélection de techniques pour tester diverses hypothèses et l'intégration innovante des signes et des symptômes dans une relation causale. Les études mécanistes commencent par l'exposition, suivent la distribution temporelle et le devenir dans le corps (pharmacocinétique) et mesurent l'effet toxique résultant à un certain niveau du système et à un certain niveau de dose. Différentes substances peuvent agir à différents niveaux du système biologique en provoquant une toxicité.
Exposition
La voie d'exposition dans les études mécanistes est généralement la même que pour l'exposition humaine. La voie est importante parce qu'il peut y avoir des effets qui se produisent localement au site d'exposition en plus des effets systémiques après que le produit chimique a été absorbé dans le sang et distribué dans tout le corps. Un exemple simple mais convaincant d'un effet local serait l'irritation et la corrosion éventuelle de la peau suite à l'application de solutions acides ou alcalines fortes conçues pour nettoyer les surfaces dures. De même, une irritation et une mort cellulaire peuvent survenir dans les cellules tapissant le nez et/ou les poumons suite à une exposition à des vapeurs ou des gaz irritants tels que les oxydes d'azote ou l'ozone. (Les deux sont des constituants de la pollution de l'air, ou smog). Suite à l'absorption d'un produit chimique dans le sang par la peau, les poumons ou le tractus gastro-intestinal, la concentration dans tout organe ou tissu est contrôlée par de nombreux facteurs qui déterminent la pharmacocinétique du produit chimique dans le corps. Le corps a la capacité d'activer et de détoxifier divers produits chimiques, comme indiqué ci-dessous.
Rôle de la pharmacocinétique dans la toxicité
La pharmacocinétique décrit les relations temporelles pour l'absorption chimique, la distribution, le métabolisme (altérations biochimiques dans le corps) et l'élimination ou l'excrétion du corps. Par rapport aux mécanismes de toxicité, ces variables pharmacocinétiques peuvent être très importantes et, dans certains cas, déterminer si la toxicité se produira ou non. Par exemple, si un matériau n'est pas absorbé en quantité suffisante, la toxicité systémique (à l'intérieur du corps) ne se produira pas. À l'inverse, un produit chimique hautement réactif qui est détoxifié rapidement (en quelques secondes ou minutes) par des enzymes digestives ou hépatiques peut ne pas avoir le temps de provoquer une toxicité. Certaines substances et mélanges halogénés polycycliques ainsi que certains métaux comme le plomb n'entraîneraient pas de toxicité significative si l'excrétion était rapide; mais l'accumulation à des niveaux suffisamment élevés détermine leur toxicité puisque l'excrétion n'est pas rapide (parfois mesurée en années). Heureusement, la plupart des produits chimiques n'ont pas une rétention aussi longue dans le corps. L'accumulation d'un matériau inoffensif n'induirait toujours pas de toxicité. Le taux d'élimination du corps et de détoxication est souvent appelé la demi-vie du produit chimique, qui est le temps nécessaire pour que 50 % du produit chimique soit excrété ou transformé en une forme non toxique.
Cependant, si un produit chimique s'accumule dans une cellule ou un organe particulier, cela peut signaler une raison d'examiner plus avant sa toxicité potentielle dans cet organe. Plus récemment, des modèles mathématiques ont été développés pour extrapoler des variables pharmacocinétiques de l'animal à l'homme. Ces modèles pharmacocinétiques sont extrêmement utiles pour générer des hypothèses et tester si l'animal expérimental peut être une bonne représentation pour l'homme. De nombreux chapitres et textes ont été écrits sur ce sujet (Gehring et al. 1976 ; Reitz et al. 1987 ; Nolan et al. 1995). Un exemple simplifié d'un modèle physiologique est illustré à la figure 1.
Figure 1. Un modèle pharmacocinétique simplifié

Différents niveaux et systèmes peuvent être affectés négativement
La toxicité peut être décrite à différents niveaux biologiques. La lésion peut être évaluée sur l'ensemble de la personne (ou de l'animal), du système organique, de la cellule ou de la molécule. Les systèmes organiques comprennent les systèmes immunitaire, respiratoire, cardiovasculaire, rénal, endocrinien, digestif, musculo-squelettique, sanguin, reproducteur et nerveux central. Certains organes clés comprennent le foie, les reins, les poumons, le cerveau, la peau, les yeux, le cœur, les testicules ou les ovaires et d'autres organes majeurs. Au niveau cellulaire/biochimique, les effets indésirables comprennent l'interférence avec la fonction normale des protéines, la fonction des récepteurs endocriniens, l'inhibition de l'énergie métabolique ou l'inhibition ou l'induction d'enzymes xénobiotiques (substances étrangères). Les effets indésirables au niveau moléculaire comprennent l'altération de la fonction normale de la transcription ADN-ARN, de la liaison spécifique aux récepteurs cytoplasmiques et nucléaires, et des gènes ou des produits géniques. En fin de compte, le dysfonctionnement d'un système d'organe majeur est probablement causé par une altération moléculaire dans une cellule cible particulière au sein de cet organe. Cependant, il n'est pas toujours possible de retracer un mécanisme jusqu'à une origine moléculaire de causalité, et ce n'est pas non plus nécessaire. L'intervention et la thérapie peuvent être conçues sans une compréhension complète de la cible moléculaire. Cependant, la connaissance du mécanisme spécifique de la toxicité augmente la valeur prédictive et la précision de l'extrapolation à d'autres produits chimiques. La figure 2 est une représentation schématique des différents niveaux où l'interférence des processus physiologiques normaux peut être détectée. Les flèches indiquent que les conséquences pour un individu peuvent être déterminées de haut en bas (exposition, pharmacocinétique à la toxicité du système/organe) ou de bas en haut (modification moléculaire, effet cellulaire/biochimique à la toxicité du système/organe).
Figure 2. Représentation des mécanismes de toxicité
Exemples de mécanismes de toxicité
Les mécanismes de toxicité peuvent être simples ou très complexes. Souvent, il existe une différence entre le type de toxicité, le mécanisme de toxicité et le niveau d'effet, selon que les effets indésirables sont dus à une seule dose aiguë élevée (comme un empoisonnement accidentel) ou à une dose plus faible. exposition répétée (due à une exposition professionnelle ou environnementale). Classiquement, à des fins de test, une dose élevée unique aiguë est administrée par intubation directe dans l'estomac d'un rongeur ou par exposition à une atmosphère de gaz ou de vapeur pendant deux à quatre heures, selon ce qui ressemble le mieux à l'exposition humaine. Les animaux sont observés pendant une période de deux semaines après l'exposition, puis les principaux organes externes et internes sont examinés pour détecter les blessures. Les tests à doses répétées varient de quelques mois à plusieurs années. Pour les espèces de rongeurs, deux ans sont considérés comme une étude chronique (durée de vie) suffisante pour évaluer la toxicité et la cancérogénicité, tandis que pour les primates non humains, deux ans seraient considérés comme une étude subchronique (moins que la durée de vie) pour évaluer la toxicité à doses répétées. Après l'exposition, un examen complet de tous les tissus, organes et fluides est effectué pour déterminer tout effet indésirable.
Mécanismes de toxicité aiguë
Les exemples suivants sont spécifiques aux effets aigus à forte dose pouvant entraîner la mort ou une incapacité grave. Cependant, dans certains cas, l'intervention entraînera des effets transitoires et entièrement réversibles. La dose ou la gravité de l'exposition déterminera le résultat.
Asphyxiants simples. Le mécanisme de toxicité des gaz inertes et de certaines autres substances non réactives est le manque d'oxygène (anoxie). Ces produits chimiques, qui causent une privation d'oxygène au système nerveux central (SNC), sont appelés asphyxiants simples. Si une personne pénètre dans un espace clos contenant de l'azote sans suffisamment d'oxygène, un appauvrissement immédiat en oxygène se produit dans le cerveau et entraîne une perte de conscience et éventuellement la mort si la personne n'est pas rapidement évacuée. Dans les cas extrêmes (proche de zéro oxygène), l'inconscience peut survenir en quelques secondes. Le sauvetage dépend d'un déplacement rapide vers un environnement oxygéné. La survie avec des lésions cérébrales irréversibles peut survenir à la suite d'un sauvetage retardé, en raison de la mort des neurones, qui ne peuvent pas se régénérer.
Asphyxiants chimiques. Le monoxyde de carbone (CO) entre en compétition avec l'oxygène pour se lier à l'hémoglobine (dans les globules rouges) et prive donc les tissus d'oxygène pour le métabolisme énergétique ; la mort cellulaire peut en résulter. L'intervention comprend l'élimination de la source de CO et le traitement à l'oxygène. L'utilisation directe de l'oxygène est basée sur l'action toxique du CO. Un autre asphyxiant chimique puissant est le cyanure. L'ion cyanure interfère avec le métabolisme cellulaire et l'utilisation de l'oxygène pour l'énergie. Le traitement au nitrite de sodium provoque une modification de l'hémoglobine des globules rouges en méthémoglobine. La méthémoglobine a une plus grande affinité de liaison avec l'ion cyanure que la cible cellulaire du cyanure. Par conséquent, la méthémoglobine lie le cyanure et éloigne le cyanure des cellules cibles. Cela constitue la base du traitement antidote.
Dépresseurs du système nerveux central (SNC). La toxicité aiguë est caractérisée par la sédation ou l'inconscience pour un certain nombre de matériaux comme les solvants qui ne sont pas réactifs ou qui sont transformés en intermédiaires réactifs. On suppose que la sédation/anesthésie est due à une interaction du solvant avec les membranes des cellules du SNC, ce qui altère leur capacité à transmettre des signaux électriques et chimiques. Alors que la sédation peut sembler une forme légère de toxicité et a été à la base du développement des premiers anesthésiques, « la dose fait toujours le poison ». Si une dose suffisante est administrée par ingestion ou inhalation, l'animal peut mourir par arrêt respiratoire. Si la mort anesthésique ne se produit pas, ce type de toxicité est généralement facilement réversible lorsque le sujet est retiré de l'environnement ou que le produit chimique est redistribué ou éliminé du corps.
Effets sur la peau. Les effets indésirables sur la peau peuvent aller de l'irritation à la corrosion, selon la substance rencontrée. Les acides forts et les solutions alcalines sont incompatibles avec les tissus vivants et sont corrosifs, provoquant des brûlures chimiques et d'éventuelles cicatrices. La cicatrisation est due à la mort des cellules cutanées profondes du derme responsables de la régénération. Des concentrations plus faibles peuvent simplement provoquer une irritation de la première couche de peau.
Un autre mécanisme toxique spécifique de la peau est celui de la sensibilisation chimique. Par exemple, la sensibilisation se produit lorsque le 2,4-dinitrochlorobenzène se lie aux protéines naturelles de la peau et que le système immunitaire reconnaît le complexe lié aux protéines altérées comme un corps étranger. En réagissant à ce corps étranger, le système immunitaire active des cellules spéciales pour éliminer le corps étranger en libérant des médiateurs (cytokines) qui provoquent une éruption cutanée ou une dermatite (voir « Immunotoxicologie »). C'est la même réaction du système immunitaire lorsque l'exposition à l'herbe à puce se produit. La sensibilisation immunitaire est très spécifique au produit chimique particulier et nécessite au moins deux expositions avant qu'une réponse ne soit déclenchée. La première exposition sensibilise (prépare les cellules à reconnaître le produit chimique) et les expositions suivantes déclenchent la réponse du système immunitaire. Le retrait du contact et le traitement symptomatique avec des crèmes anti-inflammatoires contenant des stéroïdes sont généralement efficaces pour traiter les personnes sensibilisées. Dans les cas graves ou réfractaires, un immunosuppresseur à action systémique comme la prednisone est utilisé en conjonction avec un traitement topique.
Sensibilisation pulmonaire. Une réponse de sensibilisation immunitaire est provoquée par le diisocyanate de toluène (TDI), mais le site cible est les poumons. La surexposition au TDI chez les personnes sensibles provoque un œdème pulmonaire (accumulation de liquide), une constriction bronchique et une altération de la respiration. Il s'agit d'une affection grave qui nécessite de soustraire l'individu à des expositions ultérieures potentielles. Le traitement est avant tout symptomatique. La sensibilisation de la peau et des poumons suit une dose-réponse. Le dépassement du niveau fixé pour l'exposition professionnelle peut entraîner des effets indésirables.
Effets sur les yeux. Les lésions oculaires vont du rougissement de la couche externe (rougeur de la piscine) à la formation de cataracte de la cornée jusqu'aux lésions de l'iris (partie colorée de l'œil). Des tests d'irritation oculaire sont effectués lorsqu'on pense qu'aucune blessure grave ne se produira. De nombreux mécanismes à l'origine de la corrosion cutanée peuvent également provoquer des lésions oculaires. Les matériaux corrosifs pour la peau, comme les acides forts (pH inférieur à 2) et les alcalis (pH supérieur à 11.5), ne sont pas testés dans les yeux des animaux car la plupart provoqueront la corrosion et la cécité en raison d'un mécanisme similaire à celui qui provoque la corrosion cutanée . De plus, les agents tensioactifs tels que les détergents et les tensioactifs peuvent provoquer des lésions oculaires allant de l'irritation à la corrosion. Un groupe de matériaux qui nécessite de la prudence est celui des tensioactifs chargés positivement (cationiques), qui peuvent provoquer des brûlures, une opacité permanente de la cornée et une vascularisation (formation de vaisseaux sanguins). Un autre produit chimique, le dinitrophénol, a un effet spécifique de formation de cataracte. Cela semble être lié à la concentration de ce produit chimique dans l'œil, qui est un exemple de spécificité de distribution pharmacocinétique.
Bien que la liste ci-dessus soit loin d'être exhaustive, elle est conçue pour donner au lecteur une appréciation des divers mécanismes de toxicité aiguë.
Mécanismes de toxicité subchronique et chronique
Lorsqu'ils sont administrés en une seule dose élevée, certains produits chimiques n'ont pas le même mécanisme de toxicité que lorsqu'ils sont administrés à plusieurs reprises à une dose plus faible mais toujours toxique. Lorsqu'une seule dose élevée est administrée, il y a toujours la possibilité de dépasser la capacité de la personne à détoxifier ou à excréter le produit chimique, ce qui peut entraîner une réponse toxique différente de celle obtenue lorsque des doses répétitives plus faibles sont administrées. L'alcool est un bon exemple. De fortes doses d'alcool entraînent des effets primaires sur le système nerveux central, tandis que des doses répétées plus faibles entraînent des lésions hépatiques.
Inhibition de l'anticholinestérase. La plupart des pesticides organophosphorés, par exemple, ont peu de toxicité pour les mammifères jusqu'à ce qu'ils soient activés métaboliquement, principalement dans le foie. Le principal mécanisme d'action des organophosphorés est l'inhibition de l'acétylcholinestérase (AChE) dans le cerveau et le système nerveux périphérique. L'AChE est l'enzyme normale qui termine la stimulation du neurotransmetteur acétylcholine. Une légère inhibition de l'AChE sur une période prolongée n'a pas été associée à des effets indésirables. À des niveaux d'exposition élevés, l'incapacité à mettre fin à cette stimulation neuronale entraîne une surstimulation du système nerveux cholinergique. La surstimulation cholinergique entraîne finalement une foule de symptômes, y compris un arrêt respiratoire, suivi de la mort si elle n'est pas traitée. Le traitement principal est l'administration d'atropine, qui bloque les effets de l'acétylcholine, et l'administration de chlorure de pralidoxime, qui réactive l'AChE inhibée. Par conséquent, la cause et le traitement de la toxicité des organophosphates sont abordés en comprenant la base biochimique de la toxicité.
Activation métabolique. De nombreux produits chimiques, y compris le tétrachlorure de carbone, le chloroforme, l'acétylaminofluorène, les nitrosamines et le paraquat sont métaboliquement activés en radicaux libres ou autres intermédiaires réactifs qui inhibent et interfèrent avec la fonction cellulaire normale. À des niveaux d'exposition élevés, cela entraîne la mort cellulaire (voir « Lésion cellulaire et mort cellulaire »). Alors que les interactions spécifiques et les cibles cellulaires restent inconnues, les systèmes d'organes qui ont la capacité d'activer ces produits chimiques, comme le foie, les reins et les poumons, sont tous des cibles potentielles de blessures. En effet, des cellules particulières au sein d'un organe ont une capacité plus ou moins grande à activer ou détoxifier ces intermédiaires, et cette capacité détermine la susceptibilité intracellulaire au sein d'un organe. Le métabolisme est l'une des raisons pour lesquelles une compréhension de la pharmacocinétique, qui décrit ces types de transformations ainsi que la distribution et l'élimination de ces intermédiaires, est importante pour reconnaître le mécanisme d'action de ces produits chimiques.
Mécanismes du cancer. Le cancer est une multiplicité de maladies, et bien que la compréhension de certains types de cancer s'améliore rapidement grâce aux nombreuses techniques de biologie moléculaire qui ont été développées depuis 1980, il reste encore beaucoup à apprendre. Cependant, il est clair que le développement du cancer est un processus en plusieurs étapes et que les gènes critiques sont essentiels à différents types de cancer. Des altérations de l'ADN (mutations somatiques) d'un certain nombre de ces gènes critiques peuvent entraîner une susceptibilité accrue ou des lésions cancéreuses (voir « Toxicologie génétique »). L'exposition à des produits chimiques naturels (dans les aliments cuits comme le bœuf et le poisson) ou à des produits chimiques synthétiques (comme la benzidine, utilisée comme colorant) ou à des agents physiques (lumière ultraviolette du soleil, radon du sol, rayonnement gamma provenant de procédures médicales ou d'activités industrielles) sont tous contributeurs aux mutations génétiques somatiques. Cependant, il existe des substances naturelles et synthétiques (comme les antioxydants) et des processus de réparation de l'ADN qui protègent et maintiennent l'homéostasie. Il est clair que la génétique est un facteur important dans le cancer, puisque les syndromes de maladies génétiques telles que le xeroderma pigmentosum, où il y a un manque de réparation normale de l'ADN, augmentent considérablement la susceptibilité au cancer de la peau due à l'exposition à la lumière ultraviolette du soleil.
Mécanismes de reproduction. Comme dans le cas du cancer, de nombreux mécanismes de toxicité pour la reproduction et/ou le développement sont connus, mais il reste encore beaucoup à apprendre. On sait que certains virus (comme la rubéole), des infections bactériennes et des médicaments (comme la thalidomide et la vitamine A) nuiront au développement. Récemment, les travaux de Khera (1991), examinés par Carney (1994), montrent de bonnes preuves que les effets anormaux sur le développement dans les tests sur les animaux avec l'éthylène glycol sont attribuables aux métabolites acides métaboliques maternels. Cela se produit lorsque l'éthylène glycol est métabolisé en métabolites acides, notamment l'acide glycolique et l'acide oxalique. Les effets ultérieurs sur le placenta et le fœtus semblent être dus à ce processus de toxicité métabolique.
Conclusion
Le but de cet article est de donner une perspective sur plusieurs mécanismes connus de toxicité et la nécessité d'études futures. Il est important de comprendre que les connaissances mécanistes ne sont pas absolument nécessaires pour protéger la santé humaine ou environnementale. Cette connaissance améliorera la capacité du professionnel à mieux prévoir et gérer la toxicité. Les techniques réelles utilisées pour élucider un mécanisme particulier dépendent des connaissances collectives des scientifiques et de la pensée de ceux qui prennent les décisions concernant la santé humaine.
" AVIS DE NON-RESPONSABILITÉ : L'OIT n'assume aucune responsabilité pour le contenu présenté sur ce portail Web qui est présenté dans une langue autre que l'anglais, qui est la langue utilisée pour la production initiale et l'examen par les pairs du contenu original. Certaines statistiques n'ont pas été mises à jour depuis la production de la 4ème édition de l'Encyclopédie (1998)."